





Rhawn Gabriel Joseph, Ph.D.
BrainMind.com
There are nine major somesthetic areas within the parietal lobe, such that the primary, association, and assimilation areas actually consist of numerous subareas. Broadly, and most generally, however, the parietal lobe may be subdivided into a primary receiving area (involving Brodmann's areas 3ab,1,2) within the post central gyrus, an immediately adjacent somesthetic association area (Brodmann's area 5ab), a polymodal (visual, motor, somesthetic) receiving area located in the superior-posterior parietal lobule (area 7ab), a granular insular area which is located in the inferior convexity and encompasses part of the marginal gyrus, and a multimodal-assimilation area within the inferior parietal lobule (areas 7, 39, 40) which encompasses the angular and supramarginal gyrus.
Developmentally, of all cortical regions, the inferior parietal lobule is one of the last to functionally and anatomically mature (Blinkov & Glezer, 1968; Flechsig, 1901; Conel, 1937-1943; Joseph & Gallagher, 1985, Joseph et al., 1984). Hence, many capacities mediated by this area (e.g. reading, calculation, the performance of reversible operations in space) are late to develop appearing between the ages of 5-8.
Sitting at the junction of the temporal, parietal, and occipital lobes, the inferior region (which includes the angular and supramarginal gyri) has no strict anatomical boundaries, is partly coextensive with the posterior-superior temporal gyrus, and includes part of area 7 as well as area 37. It maintains rich interconnections with the visual, auditory, and somesthetic associations areas including the middle (basal) temporal lobe, the superior colliculus via the pulvinar, the lateral geniculate nucleus of the thalamus, and massive interconnections with the frontal lobes, inferior temporal region, and other higher order assimilation areas throughout the neocortex (Bruce, Desimone & Gross, 2006; Burton & Jones, 1976; Geschwind, 1965; Jones & Powell, 1970; Seltzer & Pandya, 1978; Zeki, 1974).
THE MULTI-MODAL ASSIMILATION AREA
Over the course of evolution the amygdala, hippocampus, and medial temporal lobe began to balloon outward and upward, giving rise to superior temporal lobe, and then continuing to expand in a posterior direction, forming part of the angular and marginal gyrus. Hence, this portion of the inferior parietal lobule has auditory and thus (in the left hemisphere) language capabilities. However, with the evolution of the thumb and the capability of utilizing a precision grasp coupled with tool making and related temporal-sequential tasks, the superior parietal lobule also expanded, thereby also giving rise to inferior parietal neocortical tissue.

Given its location at the border regions of the somesthetic, auditory, and visual neocortices, and containing neurons and receiving input from these modalities, as the inferior parietal lobule evolved it became increasingly multimodally responsive; a single neuron simultaneously receiving highly processed somesthetic, visual, auditory and movement related input from the various association areas. Hence, many of the neurons in this area are multi-specialized for simultaneously analyzing auditory, somesthetic, and spatial-visual associations, and have visual receptive properties which encompass almost the entire visual field, with some cells responding to visual stimuli of almost any size, shape, or form (Bruce et al. 1982, 2006; Hyvaerinene & Shelepin, 2009).
Inferior parietal neurons are involved in the assimilation and creation of cross modal associations and act to increase the capacity for the organization, labeling and multiple categorization of sensory-motor and conceptual events (Geschwind, 1965; Joseph 1982). One can thus create visual, somesthetic, or auditory equalivalents of objects, actions, feelings, and ideas, simultaneously. For example, conceptualizing a "chair" as a word, visual object, or in regard to sensation, usage, and even price. That is, the IPL is directly involved in naming--as demonstrated by functional imaging (Price, 2017). The left IPL becomes activated when reading ( Bookheimer, et al., 1995; Menard, et al., 1996; Price, 2017 Price, et al., 1996; Vandenberghe, et al., 1996) during semantic processing (Price, 2017), and when generating words (Shaywitz, et al., 1995; Warburton, et al., 1996) or when making syllable judgements (Price, 2017). Indeed, the IPL appears to act as a pholological storehouse that becomes activated during short-term memory and word retrieval (Demonet, et al., 2004; Paulesu, et al., 1993; Price, 2017) and becomes highly active when retrieving the meaning of words during semantic processing and semantic decision tasks (Price, 2017).
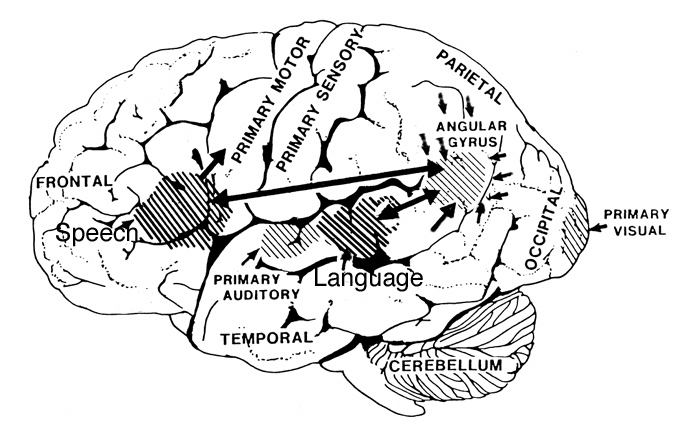
LANGUAGE CAPABILITIES
Because of it's involvement in functions such as those described above, one side-effect of damage to the left angular gyrus, is a condition called anomia, i.e. severe word finding and confrontive naming difficulty. These individuals have difficulty naming objects, describing, pictures, etc. Moreover, lesions involving the angular gyrus, or when damage occurs between the fiber pathways linking the left inferior parietal lobule with the visual cortex, there can also result Pure Word Blindness. This is due to an inability to receive visual input from the left and right visual cortex and to transmit this information to Wernicke's area so that auditory equivalents may be called up. Such patients are thus unable to read and suffer from alexia.
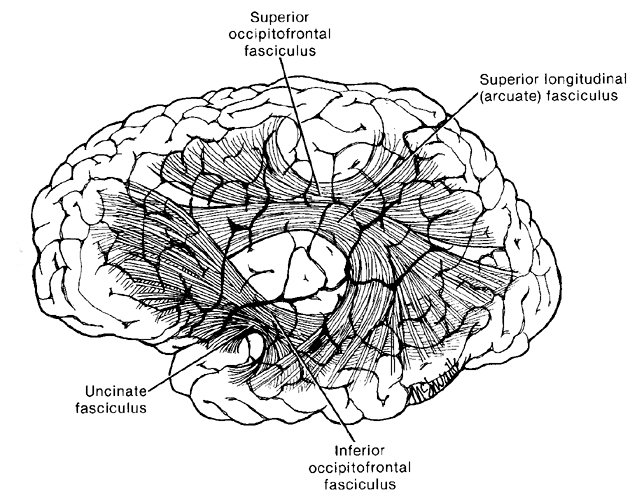
Because the inferior parietal lobule also acts as a relay center where information from Wernickes region can be transmitted, via the arcuate fasciculus, to Broca's area (for expression) destructive lesions, particularly to the supramarginal gyrus of the left cerebral hemisphere can result in conduction aphasia (see chapter 11). Although comprehension would be intact and a patient would know what she wanted to say, she would be unable to say it. Nor would she be able to repeat simple statements, read out loud, or write to dictation. This is because Broca's area is disconnected from the posterior language zones.
REFERENCES






