Rhawn Gabriel Joseph, Ph.D.
BrainMind.com
Cambrian Explosion Beginning around 540 million years ago, long after what appears to have been a mass extinction of the Ediacaran fauna, there ensued an explosion of life with all manner of complex creatures appearing in every river, ocean, and stream; an explosion of life collectively referred to as The Cambrian Explosion. These diverse and complex creatures included organisms with a hard tube-like outer-skeleton consisting of calcium carbonate, and all manner of "small shelly fish" (Anabrites, Protohertzina), as well as sponges, jelly fish, mollusks, brachipods, and the first anthropods (e.g. trilobites) which immediately sprouted legs. Thus, with no history of derivative ancestral forms, all manner of complex creatures suddenly emerged with gills, intestines, joints, and modern eyes equipped with retinas and fully modern optic lenses--optically equipped retinas indicating the presence of a well formed ganglionic brain. In fact, every phylum in existence today (including several which have since become extinct), emerged during the Cambrian Explosion, including the first representative of the phylum Chordata, i.e. creatures with a brainstem, forebrain, olfactory bulbs, and thus true brains: tunicates and the first jawless fish.
Hence, during the Cambrian epoch there was a cerebral and thus a cognitive explosion. What had been a simple network of neurons located beneath the skin, became ganglionic and concentrated in the anterior "head region" of many organisms. In some species, these ganglionic collections in turn became reorganized thereby giving rise to the the first primitive brains.
Two hundred million years after the onset of the Cambrian explosion of life most of these life forms were quite suddenly sloughed off in a mass extinction (marking the end of a period referred to as the Permian). However, this was followed by yet another explosion of life both in the sea and on dry land, the metamorphosis of which again paralleled dramatic shifts in climate, atmospheric conditions, and other environmental variables (e.g. (Butcher et al., 2012; Knoll, 2011, 2012). However, contrary to Darwinian theory, not a single new phylum emerged.
THE EVOLUTION OF THE BRAIN
Tunicates were among the first chordates (subphylum Urochordata) to evolve some 500 million years ago, and were soon followed by the first jawless fish (e.g. Astraspis, Arandaspis), who in turn gave rise to cartilaginous (Cyclostomes) "bony" fish and the sarcopterygian (lobe finned) fish. Lobed finned fish were in possession of a prototypical brain the basic framework of which would be inherited by all subsequent species, including amphibians, reptiles, and even woman and man (Nieuwenhuys & Meek, 1990b; Stephan, 1983).
Lobe finned fish were (and are) in many respects a transitional prototype for all land based creatures, as their "fleshy-lobed" fins were supported by an internal skeleton consisting of a humerus, femur, radius, ulna, tibia and fibula (Caroll, 2008; Jerison, 1973; Nieuwenhuys & Meek, 1990b; Romer, 1970). It is from these lobed fins that legs would eventually "evolve" (Caroll, 2008; Colbert, 2010; Jarvik, 2010; Jerison, 1973; Romer, 1970), and it is these lobed fins coupled with the air sacks (primitive lungs) which they had evolved, that enabled these creatures to periodically leave the water so as to venture along river banks, oceans fronts, and onto dry soil, some 400 to 350 million years ago (Caroll, 2008; Colbert, 2010; Jarvik, 2010; Jerison, 1973; Romer, 1970). Most investigators believe that it was from this group, i.e. sarcoptergia -lobed finned, and dipnoans -lung fish, that all terrestrial vertebrates evolved (Romer, 1970).
[-INSERT FIGURES 77, 78 ABOUT HERE-]
However, it was not the lobed finned lung fish but their decendants that gave rise to amphibians. These amphibian-like creatures looked something like a cross between a fish and a big salamander, with flat heads and long tails, and short stocky feet like a turtle. These include the eusthenopterons, as well as the Ichthyostegas which used four feet in order to move about (Colbert, 2010; Jarvik, 2010; Jerison, 1973; Romer, 1970). Hence, by 350 million years ago the lobe finned fish presumably evolved into a fish with legs, the eusthenopteron and ichthyostegas, which in turn evolved into amphibians, some of which grew up to 15 feet length and who sported an enlarged olfactory lobe which dominated the forebrain.
It is from one of the many branches of lobe finned fish which gave rise to amphibians who were then followed by reptiles (Caroll, 2008). With the evolutionof reptiles the dorsal pallium mushroomed in size as did the olfactory bulb, and the amygdala-striatum was pushed apart, such that the amygdala and striatum became distinct in their own right (Herrick, 1948; Nieuwenhuys, 1967). In part, this separation was induced due to the increased motor demands of living on dry land, such that the motor aspects of the striatum began to increasingly differentiate and to evolve in response to and in order to meet these new motoric needs. Yet another factor was that animals were now living in a perfumed world of smell, and these odors provided an incredible wealth of information that the forebrain became specialized to analyze. Olfactory input to the amygdala increased, thus forcing this structure to also evolve and differentiate.
Primordial reptiles split into three lineages, the anapsids which gave rise to modern turtles, synapsids which gave rise to repto-mammals and then therapsids, and diapsids which gave rise to dinosaurs, and birds, and present day reptiles (Caroll, 2008).
[-INSERT FIGURES 79, 80 & 81 ABOUT HERE-]
Repto-mammals emerged some 250 million years ago, and these creatures briefly ruled the Earth. Although the initial repto-mammals were sprawlers, the more advanced repto-mammals who in turn gave rise to therapsids, 200 million years B.P., were exceedingly technologically advanced, physically and neurologically. For example, in contrast to reptiles and amphibians, the elbows were now directed backward and the knees forward which greatly improved their ability to run and manipulate their limbs. In addition, the legs were now located beneath rather than alongside the body which enabled them to run long distances without compressing the chest and lung which allowed them to simultaneously breath while chasing prey.
Reptiles must stop in order to breath since their legs, situated alongside their body and chest cavity, constrict the expansion of the lungs as they run.
The therapsids also developed a secondary bony palate which enabled them to chew food and to simultaneously breathe without danger of choking to death. Reptiles must cease to breathe in order to swallow large chunks of their food.
Another advantage occurred in regard to thermoregulation. Therapsids became warm blooded. Reptiles must sun themselves or run around and rely on behavioral thermoregulation. For example, if a reptile fails to move from a cold to a warm location (or vice versa) their body temperature soon approaches that of the external environment. They must move about in order to gain heat by sunning themselves, or cool off by sitting in the shade.
By contrast, the hypothalamus and limbic system of the repto-mammals and then the therapsids evolved a means of regulating body temperature internally. Whereas lizards, frogs, fish, etc., have only scales, the therapsids also evolved a coat of fur, as well as sweat glands that release excessive internal heat.
Nevertheless, although exceedingly advanced, the repto-mammals were struck down and nearly became extinct (as did almost 95% of all life) following a great cataclysm when the Earth was twice struck by massive meteorites, around 250 million and 225 million years B.P. (Rampino & Haggerty, 2014), which was followed by a "giant volcanic eruption"--all of which acted to split apart the already fracture land masses and to blot out the sun with dust, thus dropping temperatures and killing off over 50% of all marine life and all larger size terrestrial animals. This gave the much smaller dinosaurs a competive advantage and the repto-mammals were displaced by these "terrible lizards."
As the environment acts on gene selection, and as the remaining repto-mammals were relegated to a nightime environment, their brain was forced to further evolve. They adapted to lurking about at night or hidden beneath deep foliage during the day--a lifestyle which induced further expansions in the olfactory dominated forebrain which became increasingly adapted to process olfactory and auditory cues. By 200 million years B.P., the repto-mammals had become therapsids (Caroll, 2008), whose dorsal pallium was now capped with a five layered (mesocortical) cingulate gyrus--the evolution of which ushered in a revolution in vocal-emotional communication and infant-maternal behavior (Joseph, 1993; MacLean, 1990).
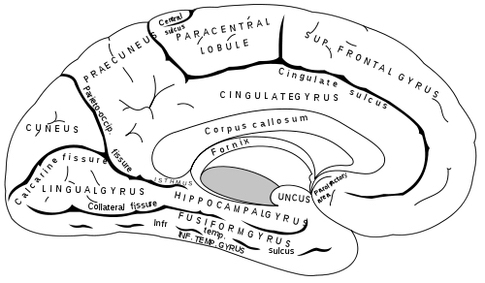
Human Brain - Medial View
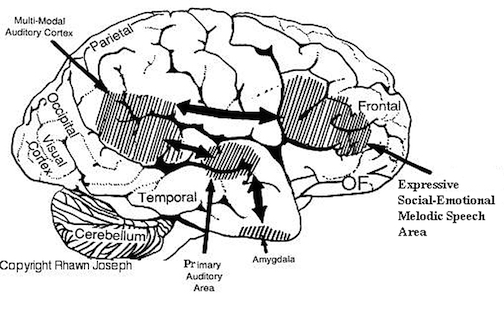
With the development of the cingulate gyrus, the therapsid's ability to communicate expanded beyond simple gestures, posturing, or olfactory-pheromonal signaling, and now included the capacity to produce a variety of complex meaningful sounds, such as between mother and infant, including, perhaps the separation cry. The five-layered cingulate gyrus provided the brain power to engage in prolonged maternal care, which in turn promoted the development of language, love, and the family (Joseph, 1993; MacLean, 1990).
As therapsids (e.g., Probainognathus from the triassic followed by Periptychus from the Paleocene), continued to evolve and to become more intelligent, the mesocortical five-layered cingulate began to sprout a small nub of neocortex (Quiroga, 2010). In later appearing therapsids, e.g. Phenocodus, the brain began to resemble that of primitive mammals, e.g., opossum or hedghog.
With the evolution of the first primitive mammals and then the more advanced mammals 130 million to 85 million years B.P., (e.g. Kumar & Hedges, 1998) this neocortical cap soon became a shroud which slowly enveloped the hemispheres of the telencephalon, and mammals soon dwarfed all previous creatures in intelligence. Slowly they began to challenge the dinosaurs for supremacy and raptor populations began to decline throughout most parts of the world with the exception of north America. However, all large sized animals, and thus most dinosaurs were given a rather sudden coupe de gra when another massive meteor struck the Yucatan peninsula, some 65 million years ago (Alvarez, 1986; Alvarez & Asaro, 1990; Rampino & Haggerty, 2014; Raup, 2011). The enormous energy released destroyed much of life in the Americas. Moreover, due to the dust thrown into the air sunlight was also blocked out for months. Temperatures dropped, thus killing off all remaining large sized cold blood animals; events which were then followed by an acid rain and a greenhouse type warming. Any remaining dinosaurs were quickly eradicated by surviving mammals, and in consequence, mammals gained dominion over the day as well as the night. As the environment acts on gene selection, the mammalian brain quickly adapted to processing visual as well as auditory stimuli, and expanded yet again.
With the ensuing evolution of primates, monkeys and apes in particular, the olfactory dominance came to be increasingly challenged and supplanted by visual and auditory input. The olfactory input to the amygdala was reduced whereas visual and auditory input became dominant. Likewise, olfactory input into the hippocampus was almost completely eliminated, being directed instead to the entorhinal cortex. Hence, the primate hippocampus and amygdala became dominated by visual and auditory input, and the entire forebrain became adapted for engaging in prolonged and detailed analysis of visual and auditory stimuli (Gloor, 1997; Stephan, 1983) as well as climbing in trees.
Over the course of later mammalian and primate evolution and as these creatures gained complete dominion over much of the planet, the neocortex began to expand at a rapid rate (Stephan & Andy, 1977). In fact, when comparing the brains of "living fossils" such as insectivores with that of humans, it appears that the neocortex expanded by a factor of 156 (even when taking into account differences in body size), whereas the 3-layered allocortex and five-layered mesocortex and all associated olfactory-limbic structures developed at a much reduced rate such that the hippocampus and septum are only 4 times larger and the amygdala is 6 times larger when comparing humans to insectivores. By contrast, the olfactory bulb is 40 times smaller (Stephan & Andy, 1969; Stephan, 1983).
Human Brain, Neocortex
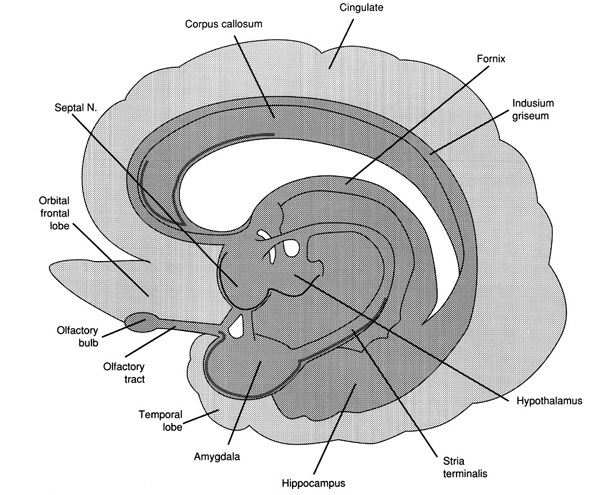
Limbic System
FROM HOMINOIDS TO HOMINIDS
It is only with the demise of the dinosaurs that primates were able to emerge from the underbrush and the darkness of night, and by 55 million years ago, during the early Eocene, at least some orders of primates (e.g. Tetonius) evolved a large occipital lobe, as well as an emerging temporal and frontal lobe (Radinksy, 1967, 1970). Thus, by 55 million years ago, ancestral primates had evolved a brain which resembled that of modern day prosimians primates.
Over the ensuing years, and as primates adapted to living in the trees, which in turn required major adaptions in the eyes and hands, the basic pattern for the primate neocortex became established and the frontal and temporal lobes continued to expand.
By 30 million years B.P., apes had emerged and these hominoids diversified and expanded their range, dwelling n Africa, China, and India, litterally from sea to shining sea. And then, from one or more of these hominoid-primate stocks, some 5 million years B.P., there emerged the first hominids who would beget Australopithecus, and H. habilis, who were followed by H. erectus--a pre-human like creature, who emerged not in Africa, but China, some 1.9 million years ago.
However, it would take another 1.4 million years before the first primitive archaic Homo sapiens would "evolve" some 500 thousands years B.P., and an additional 430 thousands years before the first H. sapiens sapiens would emerged; a peoples who were "giants" in intellect and stature, the men standing 6 feet tall on average and sporting a brain 1/3 larger than present day humans; i.e. 1800cc on average vs 1350 ccs. Over the ensuing 20,000 years, the last of the H. erectus and the last of the archaic H. sapiens would be eradicated and would disappear from the face of the Earth, presumably exterminated by the intellectually superior Cro-Magnons and other anatomically modern H. sapiens sapiens who emerged multi-regionally and spread throughout the lands.
THE CRO-MAGNONS, LANGUAGE, AND LINGUISTIC CONSCIOUSNESS
As detailed in chapter 5, it is with the evolution of the Cro-Magnon peoples, beginning at least 50,000 to 70,000 or more years ago, that we are presented with the first evidence of a fully "modern" consciousness, including language and artistic-creative capabilities equal to that of present day woman and man. These intellectual and linguistic achievements can be easily inferred based on their tools, hunting techniques, clothing, and art.
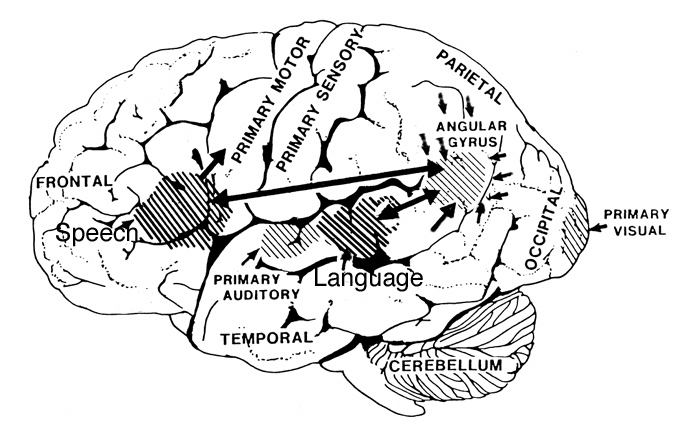
Human Brain
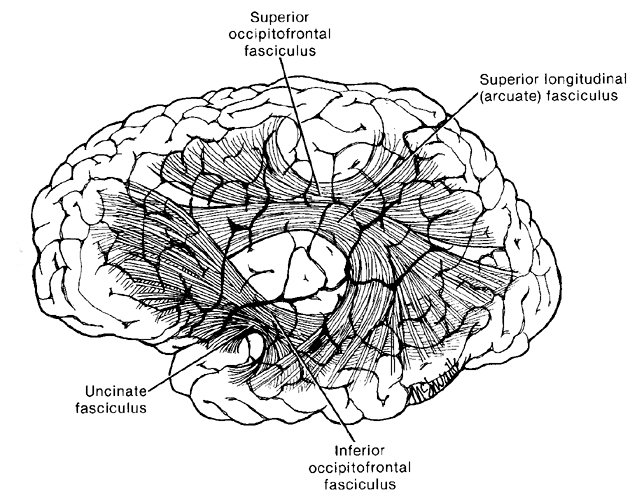
For example, with the rise of the Upper Paleolithic and the emergence of anatomically modern Upper Paleolithic human beings (e.g. the Cro-Magnon), tool making became literally and art and complex multifaceted features were incorporated in their construction and utilization. The Upper Paleolithic is characterized by the creation of complex bone and ivory tools, the sewing needle, and personal adornments such as carefully shaped beads of bone, ivory and animal teeth, animal engravings, perforated shells, statuettes, drawings, and finely crafted paintings of animal and female figures (Chauvet et al., 1996; Clark, 1967; Leroi-Gourhan, 1964; 1982; Mellars, 1989). However, since the ability to fashion and to utilize complex tools is dependent on the functional integrity of the inferior parietal lobule (Joseph, 1982, 1993; see also Barrett et al., 1998; Buxbaum, et al., 1998; De Renzi and Lucchetti, 2008; Heilman et al., 1982; Kimura, 1993; Strub and Geschwind, 1983), and as this structure is also crucial in naming and speech--as demonstrated through functional imaging (Bookheimer, et al., 1995; Demonet, et al., 2014; Menard, et al., 1996; Paulesu, et al., 1993; Price, 1997; Vandenberghe, et al., 1996)--it can thus be assumed that with the evolution of the Cro-Magnon, these people had acquired the capability of producing complex, vocabularly-rich, speech (Joseph, 1999e).
By contrast, whereas perhaps up to 90% of the Cro-magnon were right handed, and were capable of making complex multi-faceted tools, only perhaps up to 60% of Australopithecines favored the right extremity (Dart 1953), and any tools they wielded could have been no more complex than simple stones. By 1.6 million years ago, perhaps as many as 70% of H. habilis had developed similar right handed inclinations (Toth 1985). However, the stone tools associated with these people, were also exceedingly primitive, and had remained unchanged since their inception, 2.8 million years ago.
The tool making tradition of H. habilis is referred to as Oldowan. These stone tools were made by direct percussion of striking two rocks together, thus forming, for example, a rock with a cutting edge. This remained the level of sophistication until around 1.5 million years ago when the Acheulean hand ax and H. erectus appeared. The Acheulean tradition which included stone choppers and cleavers, remained the dominant technology for another million years, until around 130,000 B.P., when it was supplemented by the development of the Mousterian technology. The Mousterian is associated with the Middle Paleolithic and archaic H. sapiens and included a number of single purpose stone tools, some of which were hafted onto wooden handles. Perhaps up to 80% of archaic humans were right handed (Cornford 1986).
It thus appears that tool making capabilities paralleled the development of handedness--both of which reflect upon the functional development of the IPL. Hence, it is not until emergence of the Cro-Magnon that we have clear evidence for the complete evolution of the IPL (angular and supramarginal gyrus), and thus evidence for the temporal sequential and neurological foundations for word rich, grammatically complex, modern human language (Joseph, 1993, 1999e,f). Indeed, the IPL (i.e. angular gyrus) is unique to humans, and is crucially evolved in controlling temporal sequential hand movements including the manipulation of external objects and internal impressions (De Renzi & Lucchetti, 2008; Heilman et al., 1982; Kimura, 1993; Strub & Geschwind, 1983). In fact, the motor engrams that make possible temporal and sequential motor acts, including those involved in grammatical verbal expression, are partly localized within the IPL (De Renzi and Lucchetti, 2008; Heilman et al., 1982; Kimura, 1993; Strub and Geschwind, 1983). As noted, the IPL not only interacts with but appears to program the frontal motor areas for the purposes of producing fine motor, temporal-sequential movements, including the vocalization of speech units.

Specifically it appears that over the course of evolution, with the development of right handedness and across gathering/tool-making generations, and as the environment acts on gene selection, the IPL/angular gyrus emerged as an extension of the auditory area in the temporal lobe and the superior parietal visual-hand area which contains neurons which guide hand movements and which respond to visual input from the periphery and lower visual fields (Motter & Mountcastle, 1981; Previc 1990); i.e. the visual areas in which tool-making hands are most likely to come into view.
Because most individuals would use the right hand for tool making and the left for holding the tool, it is the left parietal lobe (which monitors the right lower visual field and controls the right hand) that guides and visually observes, learns and memorizes hand-movements when gathering, gesturing, or manipulating some object or constructing a tool (Joseph, 1993, 1999e). Over the course of evolution and as experience and the environment act on gene selection and induce neural plasticity, the parietal (and superior temporal) lobe expanded as did the IPL, and neuroplastic alterations were induced in the adjoining motor-hand area in the frontal lobe including what would become Broca's speech area.
As Wernicke's area, the parietal-hand areas and the IPL expanded, auditory input began to be sequenced, and Wernicke's area became specialized for perceiving and comprehending language units. In addition, the arcuate fasciculus axonal pathways leading from the IPL to Broca's areas also significantly expanded and increased in density, and Wernicke's area became tightly linked with and began transmitting auditory-linguistic signals to Broca's area thus inducing neurplastic alterations in these tissues (Joseph, 1993, 1999e). Hence, Wernicke's area and the IPL began sequencing auditory input, and Broca's area was transformed from a hand area to a speech area and ceased to control hand movements. Instead Broca's area began to organize the adjacent primary motor oral-laryngeal areas so as to express the words and sentences transmitted from the IPL and Wernicke's area.
Once these neural-plastic transformation took place, "limbic language" (emotional speech) became hierarchically represented, yoked to the neocortex and subject to fractionization, temporal sequencing, and multi-classification. Wernicke's area was now able to communicate with Broca's area, with the IPL injecting temporal sequences, assimilated associations, and names and words into the stream of language and thought. Hence, in addition to manipulating tools in a temporal sequential fashion, the evolution of the IPL/angular gyrus in conjunction with Broca's area and the hand-area in the frontal lobe, enabled humans to manipulate the internal environment and to transmit linguistic impulses to the frontal motor areas controlling the oral-laryngeal musculature, thereby reorganizing Broca's area in order to vocalize units of speech and giving birth to linguistic consciousness.
As these events appear to have coincided with the emergence of the Cro-Magnon, it thus appears that the cerebral hemispheres probably also became fully functionally specialized and thus lateralized as is common among modern humans. This is because, with the evolution of language and linguistic consciousness, non-verbal functions formerly associated with the left hemisphere were displaced due to functional crowding, such that the left half of the brain became dominant for language and temporal-sequential analytic activities, whereas the right hemisphere became dominant for visual spatial, affective, and non-verbal intellectual functions including maintenance of the body image. In consequence, the neocortex and the cerebral hemispheres became organized such that in addition to the mental systems associated with the brainstem and limbic system, two potentially independent forms of conscious awareness emerged and came to coexist, literally side by side.