





Rhawn Gabriel Joseph, Ph.D.
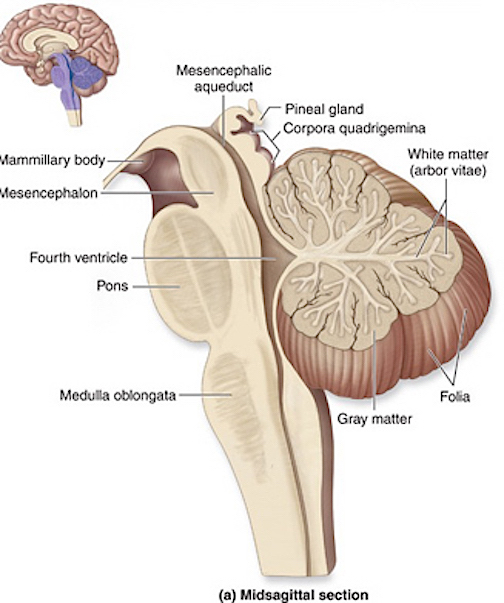
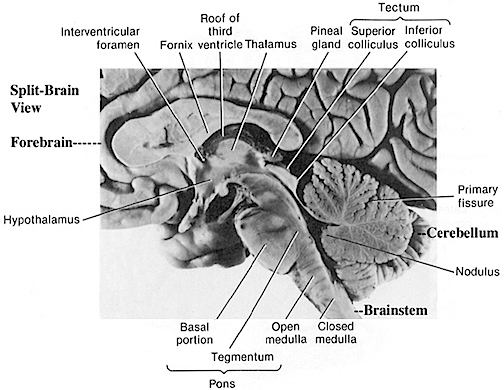
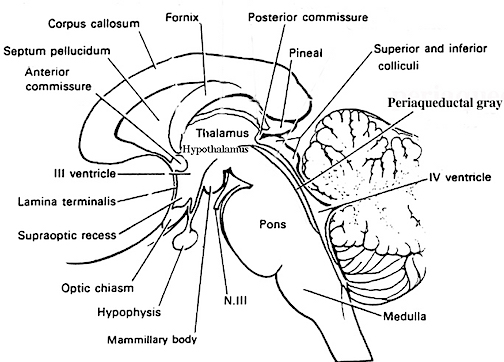
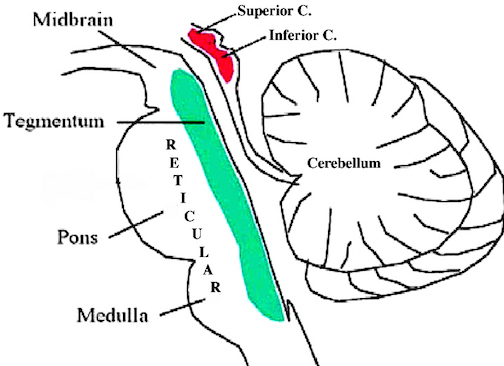
The brainstem consists of the medulla (myelencephalon), the pons (metencephalon), and the midbrain (mesencephalon). Straddling the dorsal surface of the pons is the cerebellum and spanning the length of the brainstem are the nuclei for cranial nerves 3-12.
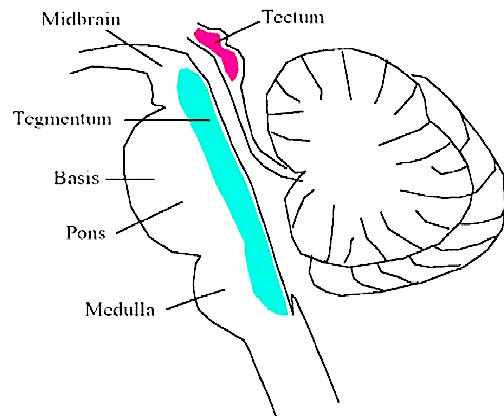
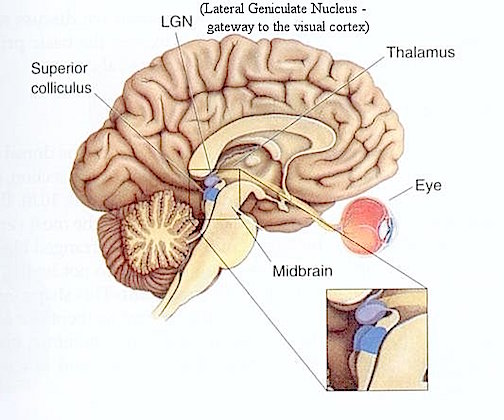
The most rostral portion of the brainstem, i.e. the midbrain merges with the diencephalon which in turn is surrounded by the cerebral hemispheres. The more posterior portion of the midbrain consists of the superior (visual) and inferior (auditory) colliculi which are directly related to the thalamus, and the red nucleus which receives motor fibers from the cerebellum and frontal and parietal lobe. The dorsal roof of the midbrain is referred to as the tegmentum (which produces dopamine) whereas more centrally located is the substantia nigra, a major source of corpus striatal dopamine.
Spanning the medial and lateral lengths of the brainstem and midbrain is a complex reticulum of richly interconnected cells with long ascending and descending axons, collectively referred to as the reticular activating system, which is concerned with generalized and selective arousal and activation of the neuroaxis. The pontine tegmentum and the more dorsal portion of the midbrain tegmentum is also considered part of the reticular formation through which course ascending sensory fibers and descending motor fibers.
MIDBRAIN EVOLUTION
The brainstem (excluding the midbrain) may have evolved in a posterior-caudal and in an anterior direction so as to merge with the visual (midbrain) areas that probably initially supplied photopic derived energy to run the motor system and to arouse and energize the brain. In this regard, evolution parallels ontogeny (see chapter 23). As also detailed in chapter 5, the anterior most segment of the midbrain and anterior pons, appear to have evolved from sensory-motor, and photosensitive cells.
Due presumably in part to its photosensitive origins, and thus its concomitant ability to extract energy, the midbrain-pontine region of the brainstem developed the capacity to respond to light vs darkness, and to manufacture various neurotransmitters; e.g. serotonin, dopamine, and norepinephrine. In this manner the brainstem is able to exert widespread influences on cerebral arousal and neural transmission.
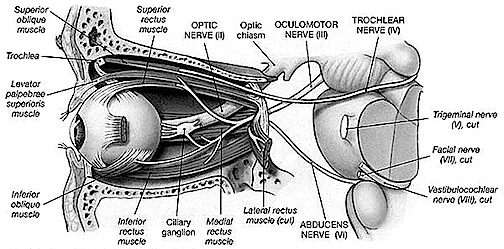
Over the course of evolutionary development, the capacity to respond to visual input increased beyond detecting light vs shadow, and soon came to include motion, as well as the ability to rotate the eyeballs and focus the retina via cranial nerves 2, 3, 4, 6. These complex functions in turn became subject to the control of the superior colliculus (or tectum).
However, with the evolution of "hearing" the midbrain became increasingly modified to subserve complex auditory functioning as well, which in turn became associated with the evolution of the inferior (auditory) colliculus. In this regard, just as the control and governance of motor functioning becomes more complex and hierarchically (caudal to anterior) organized, so too does sensory analysis and related sensory-motor integrative functions which take place within the midbrain. Multi-modal processing and complex behavioral reactions, and the integration and analysis of auditory, visual-tactile, and even motor stimuli are characteristics of the visual (superior) and auditory (inferior) colliculi of the midbrain.
THE MIDBRAIN SUPERIOR & INFERIOR COLLICULI
THE SUPERIOR COLLICULUS The superior colliculi are located in the anterior half of the midbrain. Although the midbrain per se is not well differentiated, these nuclei consist of gray and white layers and resemble cerebral cortex (Carpenter 2008). As noted in chapter 5, neocortical layers I and VII (between which are sandwiched layers II-VIa), in turn appear to be related to and/or an outgrowth of the midbrain.
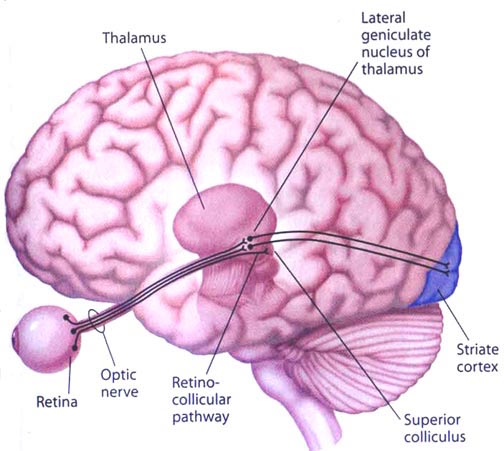
The superior colliculi can be functionally divided into superficial and deep layers which subserve somewhat different functions. (Cowie & Robinson 2009; Davidson & Bender 2008; Weller 1989). For example, the superficial layers receive considerable input from the retina as well as temporal and occipital visual cortex, and respond to moving stimuli. The superficial layers also project to vision related cranial nerve nuclei.
By contrast, the intermediate and deeper layers receive converging motor, somesthetic, auditory, visual and reticular input, and in fact serves as an extension of the reticular formation, and maintains interconnections with the caudal medulla and those cranial nerves associated with movement of the head. Hence, these layers serve as a multi-modal assimilation area which is concerned with orienting toward external stimuli and movement; particularly movements of the head and eyes during gaze shifts (Cowie & Robinson 2009; Davidson & Bender 2008; Keller & Edelman 2009).
The deep and superficial portions are also intimately interconnected (Carpenter, 2008; Parent 1995) and both receive extensive optic input from the contralateral eye, and contralateral visual input from the ipsilateral eye. Moreover, they receive ipsilateral projections from the neocortical visual areas regarding the contralateral half of visual space. Hence, each colliculus is concerned with either the right or left half; i.e. the contralateral half of visual space. In addition, the colliculi receive input from the frontal eye fields.
As noted, cells within the colliculi are responsive to visual motion and many tend to respond only to movement in a certain direction (Davidson & Bender 2008). Most do not respond to stationary stimuli. Given that reptiles, amphibians, and fish are devoid of higher cortical centers, it thus appears that the colliculi evolved so as to detect the presence of prey (or predators) and to guide orienting reactions and thus movement related to escape or food procurement.
THE INFERIOR COLLICULUS
The inferior colliculus (IC) is predominantly concerned with detecting and analyzing auditory stimuli, and in fact is tonotopically organized; i.e. neurons are arranged in a laminar pattern which represents different auditory frequency bands (reviewed in Carpenter 2008; Brodal 1981). The IC can also respond to sounds arriving from either ear. This enables the IC to analyze and localize the source of various sounds and to correlate them with their various spatiotemporal characteristics. In this manner, sounds can be identified as coming from a certain direction and from a specific source.
The IC sends fibers to the pons and medulla as well as the superior colliculus, and relays auditory impulses to the spinal cord, and the nuclei subserving the neck and facial musculature (Henkel & Edwards, 1978; Wright & Barnes, 1972). Hence, via the IC, auditory impulses can trigger head and body turning and thus orientation toward sound sources. In mammals, the IC also acts to relay auditory signals received from the lateral lemniscus to the medial (auditory) geniculate of the thalamus, and in this regard it serves not only to analyze and orient toward auditory stimuli, but as a major auditory relay nucleus. However, in this regard it is completely dependent on cranial nerve nuclei subserving the vestibular-cochlear system; cranial nerve VIII which is located at the pontine, medulla junction.
BRAINSTEM AUDITORY PERCEPTION
Long before amphibians left the sea to take up life on land and in the air, fish had already learned to detect and analyze sounds made in the water. This was made possible by a structure located along both sides of the fish's body, called the lateral line. The lateral line is very sensitive to vibrations including those made by sound. The mammalian auditory system, however, did not evolve from the lateral line but from the vestibular nucleus (chapter 5).
The human brainstem vestibular nucleus is very sensitive to vibrations and gravitational influences and helps to mediates balance and movement, such as in swimming and running. Over the course of evolution, it is through the vestibular nucleus (and the amygdala) where the first neuronal rudiments of "hearing" and the analysis of sound also took place. Among all mammals this is a function the vestibular nuclei continues to serve via the signals it receives from the inner ear; the brain's outpost for detecting certain vibratory waves of molecules and their frequency of occurrence.
Initially, however, the auditory system also evolved so that organisms could turn and orient toward and localize unexpected sound sources. This was made possible by messages transmitted to the midbrain inferior colliculus. This includes things that go bump in the night such as the settling of the house and the creaking of the floor, all of which can give rise to alarm reactions in the listener (in their amygdala). This is a sense humans and their ancestors have maintained for well over several million years, its main purpose being to maximize survival via the detection of approaching predators. In fact, the inferior colliculus (as well as the amygdala of the limbic system) was the main source of auditory analysis two hundred million years before the evolution of neocortex (see chapter 5). This is a function it continues to perform among modern day reptiles, amphibians, sharks, as well as mammals.
AUDITORY TRANSMISSION FROM THE COCHLEA TO THE TEMPORAL LOBE
Within the cochlea of the inner ear are tiny hair cells which serve as sensory receptors. These cells give rise to axons which form the cochlear division of the 8th cranial nerve; i.e. the auditory nerve. This rope of fibers exits the inner ear and travels to and terminates in the cochlear nucleus which overlaps and is located immediately adjacent to the vestibular-nucleus from which it evolved within the brainstem.
Among mammals the cochlear nucleus in turn projects auditory information to three different collections of nuclei. These are the superior olivary complex, the nucleus of the lateral lemniscus, and, as noted, the inferior colliculi.
A considerable degree of information analysis occurs in each of these areas before being relayed to yet other brain regions such as the amygdala (which extracts those features which are emotionally or motivationally significant), and to the thalamus where yet further analysis occurs.
Among mammals, from the thalamus and amygdala, auditory signals are transmitted to the primary auditory receiving area located in the neocortex of the superior temporal lobe; Heschl's gyrus. Here auditory signals undergo extensive analysis and reanalysis and simple associations begin to be formed. However, by time it has reached the neocortex, auditory signals have undergone extensive analysis by the thalamus, amygdala, and the other ancient structures mentioned above.







DVD - Brain Lectures
Six Lectures
3 DVD set = $29.95(+ shipping)
Pay By Paypal.com
DVD 1: Brain Overview
DVD 2: The Left Hemisphere, Brainstem, Midbrain, Thalamus
DVD 3: The Frontal Lobes: Frontal Lobotomy, Memory, Aphasia, Paralysis
DVD 4: The Parietal Lobes: Touch, Body-in-Space, Body Image, Hemi-Neglect, Phantom Limbs,
DVD 5: The Temporal Lobes: Language, Memory, Aphasia, Hallucinations, Face Recognition
DVD 6: The Limbic System: Amygdala, Hippocampus, Hypothalamus, Sex, Emotion, Memory, Stress, PTSD, Hallucinations