





Rhawn Gabriel Joseph, Ph.D.
BranMind.com
Sex Differences Are Pervasive One need only gaze at the human male vs female body to be presented with obviously profound differences between the sexes. Female's are less muscular, their hips are wider, and they have a vagina, uterus, fallopian tubes, a menstrual cycle, and can become pregnant and have babies, whereas, on average, males are taller, have greater upper body strength, facial hair, a penis, and are decidedly more aggressive and violent. Human males and females differ genetically, biologically, physically, hormonally, neurologically, psychologically and cognitively including in the realms of language and visual-spatial skills; all of which contribute to sex differences in consciousness.
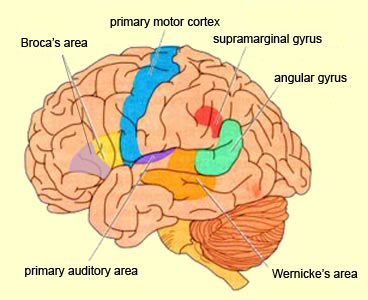
Consciousness & Language Consciousness, like cognition, is a multiplicity of mind, one aspect of which relies upon language (Joseph 2010, 1982, 2009, 2011a). Thus we often think-in-words. Thinking-in-words is language internalized, one area of the brain and mind producing these word-thoughts, another region comprehending them. These language-related tissues of the mind correspond to Broca's expressive speech area which serves as the final common pathway where thoughts are organized and expressed or experienced as words, and Wernicke's receptive speech area which comprehends these word-thoughts, be they spoken aloud or experienced in the privacy of one's own head (Joseph 2010, 1982, 1988, 20011a; Joseph et al., 1984).
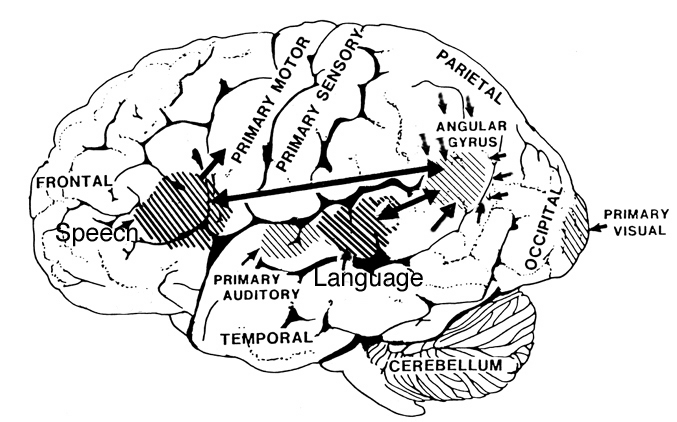
Linking Wernicke's and Broca's areas is the angular gyrus of the inferior parietal lobule which contribute to language comprehension and expression and the assimilation of multi-modal concepts from multiple modalities which enables the mind to create multiple multi-sensory categories for different words. Thus, for example, the word mustang can be conceived of as a horse, different colors of a horse, a horse running or galloping, or as a car of varying shapes, colors, and sizes.
As human language, by definition, is unique to humans, the evolution of language, and thus those aspects of the brain and mind which produce and comprehend language, have given rise to a form of consciousness which is also unique to humans, i.e. human consciousness. However, not only is human consciousness a multiplicity with different mental realms concerned with different aspects of conscious experience, but the multiplicity of the mind and thus human consciousness and cognition differs depending on if one is a man or a woman and which areas of the brain is being used to support or provide content to consciousness. In fact, non-human animal consciousness and cognition also differ depending on if that animal is male or female; and these gender differences, at least in the realm of language, have become more pronounced with the evolution of humans.
Sex Differences in Other Species There have been numerous research reports examining the biological, psychological, cultural, and political influences which are presumed to influence and shape sex differences in cognition, emotion, language, and human sexual behavior. Certainly, these and other environmental factors can have a significant impact on the brain and mind (Joseph 1982, 1992a, 1996, 1998, 1999ab). However, some of the same sex-specific cognitive traits and sex differences in sexual behavior, are demonstrated by other species (Joseph, 1979, Joseph et al. 1978; Joseph & Gallagher 2010) which evolved long before humans (Joseph 1992a,b, 1993, 1996, 1999a). These later findings demonstrate that many sex differences are biological. However, some of these sex differences became exaggerated in humans following the emergence of language, right hand dominance, and the capacity to create and utilize complex tools.
Language & Right Handedness Language is directly related to human handedness, which is explains why the areas subserving the hand and mouth are adjacent to each other in the frontal-parietal sensory motor areas, and why most humans tend to move the right hand when speaking and use the right hand for writing. However, although the ability to vocalize is characteristic of almost all animal species, handedness, it is only humans who predominantly show right hand dominance; in among approximately 90% of the population. Human language is also grammatical, and organized, expressed and comprehended in a rapid, temporal sequential fashion. As language and handedness are both subserved by adjacent areas in the left hemisphere of the human brain, it thus appears that the evolution of language is directly related to handedness and the ability to engage in rapid, temporal sequential fine motor movements of the hands and fingers all of which are associated with the functional integrity of the left hemisphere in the majority of the population. In consequence, language is temporal-sequential and grammatical, most people move their right hand and arm when talking, and use the right hand for writing (Joseph 1982, 1988a, 1990a, 1996a,b, 2000a).
These capacities and cognitive abilities are related to ancestral domestic activities traditionally associated with women, such as gathering, food preparation, the fashioning and cleaning of clothing, and prolonged child care which promotes mother-infant mutual vocalization (Joseph 1992, 1993, 1996, 2000a,b). Since the environment acts on gene selection and neuroplasticity over the course of human evolution, as these new abilities developed, the motor areas and neocortex of the left hemisphere of both males and females became functionally reorganized. Older cognitive functions associated with the left hemisphere were crowded out and displaced due to competition for synaptic space, and the left hemisphere became dominant for handedness and language in both sexes. However, as tasks traditionally associated with females not only promoted speech and language development but bi-manual fine motor activity of both hands, the female brain became bilaterally organized for language and fine motor tasks.
SEX DIFFERENCES IN LANGUAGE & VISUAL SPATIAL SKILLS
It has been consistently demonstrated that human females demonstrate superiorities on tasks involving fine motor movements of the fingers and hands (Broverman et al., 1968; Hampson and Kimura, 1992; Junaid and Fellowes 2006; Larson et al. 2007; Thomas and French 1985). Female superiorities have also been repeatedly demonstrated across a variety of verbal-measures of language ability (Deary, 2007; Johnson & Bouchard 2007; Strand et al. 2006) including and especially reading and writing (Bae et al., 2000; Hedges and Nowell 1995; Willingham and Cole 1997; Strand et al. 2006). The female advantage in verbal abilities is cross cultural, international, and is evident by age 2 (Mullis et al., 2003; National Center for Education Statistics, 2002; Ogle et al., 2003), whereas the greater propensity for females to engage in rapid movements of the upper extremities is evident even in rats (Joseph & Gallagher, 2010).



Language & Competition for Synaptic Space As tasks traditionally associated with females not only promoted speech and language development but bi-manual fine motor activity of both hands the female brain is bilaterally organized for language (Shaywitz, et al., 1995) and for performing fine motor tasks (Broverman et al., 1968; Hampson and Kimura, 1992). In consequence, ancestral visual-spatial functions associated with the right and left hemisphere were crowded out of the female brain and diminished due to competition for synaptic space. The same is not true for males. Thus, by contrast, males demonstrate inferior verbal skills and superior visual spatial and gross motor skills and this dominance also became more pronounced over the course of human evolution.
Competition for synaptic representation in the primate brain was first demonstrated in 1978 (Casagrande & Joseph 1978, 2010; Joseph & Casagrande 1978, 2010). Specifically, primates were reared with one eye-lid sutured shut, allowing only non-pattern illumination to be transmitted from the effect eye to the lateral geniculate nucleus of the thalamus (which subserved vision) and the visual neocortex. When the sutures were removed, all animals were blind in the effected eye, and synaptic space normally occupied by nerves from that eye had been hijacked by neurons from the normal eye. Neurons from the eye which was not being used, had lost the competition for synaptic space. However, when the normal eye was removed, the struggle for synaptic representation was reversed, and the formally deprived eye reestablished synaptic connections and regained vision (Casagrande & Joseph 1978, 2010; Joseph & Casagrande 1978, 2010).
Likewise, it was subsequently determined that following brain damage in humans, and the severing of axonal-neuronal interconnections, that normal neurons closest to the brain areas which had been vacated established synaptic connections with these tissues. For example, patients who had suffered parietal lobe injuries--an area of the brain which subserves the body image--experienced altered synaptic representation, such that if the hand or arm were stimulated, they instead experienced the sensation on another body part whose neurons took over the temporarily vacated synaptic space (Joseph 1986a, 1988a, 1990, 1996). Thus, there is a continual struggle and competition for synaptic representation in the human brain. Therefore, over the course neuronal representation for various cognitive functions also came to be altered, such that ancestral sex differences in cognition came to be exaggerated and other functions displaced with the evolution of handedness and language.
Hunting & Spatial Abilities Over the course of human evolutionary history, males were the hunters. Hunters require good visual spatial directional skills so as to find their way over long distances. A good hunter must be able to kill with accuracy by throwing spears or firing arrows at an animal which may be running or standing some distance away. Moreover, hunting does not promote nor does it require language as is evident in all species which hunt. Among humans, a talkative hunter will scare off all game. Thus we see that human males out-perform females on visual-spatial tasks (Hugdahl et al., 2006; Irwing & Lynn 2005; Johnson & Bouchard 2007) including accuracy in throwing (Joseph 1992a, 1993).
Male superiorities on spatial measures are evident by age 5 ( Levine et al., 1999), and include mental rotation, 3-dimensional thinking, maze learning, and the understanding of maps and directional abilities (Brown et al., 1998; Cutmore et al., 2000; Loring-Meier & Halpern, 1999; Masters & Sanders, 1993; Nordvik & Amponsah, 1998). Male spatial learning superiorities been also demonstrated in other species including rats (Joseph 1979, Joseph & Gallagher, 2010; Joseph et al., 1978).
COGNITION AND SEX DIFFERENCES IN OTHER SPECIES
The fact that these sex differences are evident in other species and can be reversed by hormonal manipulations (Joseph 1979, Joseph & Gallagher, 2010; Joseph et al., 1978), completely disproves the conspiracy theorists who argue that nurture, rearing habits, sex-steoyping, and discrimination are the responsible agents.
Specifically, Joseph (1979; Joseph & Gallagher, 2010) demonstrated when rats were raised in an enriched environment vs a deprived and restricted environment, that male rats consistently demonstrated a highly significant superiority over females in spatial maze learning. Further, even when all animals learned the spatial tasks, males demonstrated superior retention of this information and superior long-term memory for spatial details. Thus, when provided identical experiential opportunities beginning in infancy, and regardless of these environments, these sex differences remain including a propensity for females to persist in making rapid movements of the upper extremities (Joseph & Gallagher, 2010).
This also explains why, despite the fact that both sexes have experience with drinking from glass containers, males outperform females when asked to specify the horizontal water level in a tilted glass (Morris, 1971; Thomas et al., 1973). That is, females tend to erroneously draw the water as tilted when the glass is tilted, whereas males will draw the water as horizontal.
Male rats are also significantly more exploratory and show a greater propensity to investigate their environment even when both males and females are raised in an enriched environment (Joseph & Gallagher, 2010). In fact, human male children also demonstrate superior spatial abilities, and are also far more likely than females to explore a strange environment.
Moreover, Joseph et al. (1978) found that these sex differences could be reversed, biologically, through the administration of the "male" hormone, testosterone, to females, or by preventing the release of testosterone in males, during the time period when the brain is sexually differentiating. For example, female rats exposed to testosterone during early brain development when the brain is sexually differentiating, perform similarly to males on maze learning tasks, whereas castrated-infant males later perform similarly to females (Joseph et al., 1978). Likewise, homosexual men (whose brains, in some cases, may have been feminized prenatally, e.g. Meyer-Bahlburg, 1993) tend to perform more poorly than heterosexual men on spatial tasks and similar to females on verbal tasks (Gladue et al., 1990; however see Gladue & Baily, 1995). Moreover, human and non-human females exposed to high levels of masculinizing hormones and androgens during early brain development demonstrate superior visual-spatial skills, and are more competitive and aggressive as compared to normal females (Money and Ehrhardt, 1972; Ehrhardt and Baker, 1974; Joseph et al., 1978; Reinisch and Sanders, 1992; Mitchell, 1979). Hunting and the killing of prey is related to visual-spatial perceptual functioning and the tendency to behave in an aggressive fashion.
Sex Differences in Brain & Behavior Clearly these and other sex differences are biological as they are present in early childhood and in species who evolved before humans. Further, sex differences have been well documented in the structure and organization of the human and non-human brain, the limbic system in particular (Allen et al. 1989; Bleier et al. 1982; Bubenik & Brown, 1973; Filipek, et al., 1994; Nishizuka & Arai, 2011; Rainbow et al. 1982; Raisman & Field, 1971, 1973), and including those tissues of the mind which mediate male vs female sexuality, emotion, language, and spatial skills (Joseph et al., 1978; Hart et al., 1985; Lisk, 2007, 1971; Maclean, 1973; Robinson and Mishkin, 1968; Stoffels et al., 2010). For example, because a heterosexual male possess a "male" brain, he responds with sexual arousal to the female body, is more aggressive and violent, and demonstrates superior visual-spatial skills. By contrast, because the heterosexual female possesses a brain organized in the "female" pattern, she demonstrates superior language skills, is more social and emotional and nurturing, and desires to have babies and to hold and talk to babies. The brain is part of the body and it is precisely because she has a "female" brain and body that women have a menstrual cycle, become pregnant, have babies, and nurse those babies.
Moreover, it is precisely because human males are more aggressive and physically violent (which makes them good hunters) whereas females are biologically designed to produce and nurture helpless big brain babies and to talk to these babies while engaged in domestics tasks (such as food preparation, gathering) which can be performed close to the home base, that some of the sex differences apparent in other species have become exaggerated over the course of human evolution.
Evolution of Hunters & Gatherers As first proposed and theorized by Joseph in 1992, sex difference in spatial abilities and vocalization became exaggerated in humans with the evolution of "the big brain" and full-time sexual receptivity in females during the later stages of Homo erectus evolution, followed by the emergence of fine motor skills and language with the evolution of Upper Paleolithic humans, e.g. the Cro-Magnon. Although the evolutionary history of human sex differences can be traced back over 5 million years and to other species (Joseph 2000a,b 2011) the great divide in male vs female cognition and consciousness may have had had its onset about 500,000 years ago and began to accelerate around 35,000 years ago with the evolution of modern human language, complex tool use and construction, and a full time division of labor between the sexes. This "hunter-gatherer" theory of sex differences in behavior, language, cognition, and consciousness has subsequently been adopted by other scientists.
HOMO ERECTUS: SEXUALITY AND THE HOME BASE
There is no general agreement as to the various phylogentic relationships shared by the wide variety of Plio-pleistocene hominids so far discovered, and it is not yet established if present-day humans descended from Australopithecus, Homo habilis, or both or neither. Nevertheless, following and overlapping with H. habilis and Australopithecus were a wide range of quite different individuals collectively referred to as Homo erectus.
Homo erectus were big and robust, with thick browridges, large teeth, and bulging shoulder muscles (Day, 1996; Potts, 1996; Rightmire, 1990). This diverse species of humanity ranged throughout Africa, Europe, Russia, Indonesia and China from approximately 1.9 million until about 300,000 years ago, with a few isolated populations possibly hanging on in the island of Java, until 27,000 years B.P. (Day, 1996; Potts, 1984, 1996; Rightmire, 1990; Swisher et al., 1996). Presumably, H. erectus is the common ancestor for Neanderthals, the Cro-Magnon, and modern humans. However, it is just as likely that modern humans have descended from different branches of the species referred to collectively as "H. erectus".
About 1.5 million years ago H. erectus learned to harness fire and by 500,000 B.P., the first hearths began to appear in China, France, Hungary and elsewhere (Clark & Harris 1985; Isacc, 1982; Rightmire, 1990). By 500,000 B.P. H. erectus was regularly constructing crude shelters and home bases (Clark and Harris 1985; Potts 1984, 1996; Rightmire, 1990; Zhang, 1985); achievements that may have coincided with a major change in female sexuality.
BREASTS, BUTTOCKS & COSMETICS
About 1.5 million years ago H. erectus learned to harness fire and by 500,000 B.P., the first hearths began to appear in China, France, Hungary and elsewhere (Clark & Harris 1985; Isacc, 1982; Rightmire, 1990). By 500,000 B.P. H. erectus was regularly constructing crude shelters and home bases (Clark and Harris 1985; Potts 1984, 1996; Rightmire, 1990; Zhang, 1985); achievements that coincided with a major change in female sexuality (Joseph 2000a,b, 2011).
For example, during the evolutionary transition which led from Australopithecus and H. erectus and full time bipedalism, the vaginal canal underwent a reorientation from back to front, which enabled males and females to face one another during sexual intercourse, thus promoting interpersonal intimacy. With the exception of the Bonobo who are more variable, all other primates and non-human animals generally assume a dorsal ventral posture when mating (Ford and Beach, 1951; Goodall, 1986, 1990, 2010; Wickler, 1973).
This vaginal reorientation may have coincided with an expansion in the size of the brain, alterations in the size of H. erectus pelvis, and the emergence of full time female sexual receptivity. That is, with the evolution of H. erectus the female estrus cycle and the hominoid menstrual cycle which generally dictates sexually receptivity, may have undergone a transition such that, unlike all other (non-captive) female species, but like modern day woman, the H. erectus female became sexually receptive at all times and evolved enlarged breasts and buttocks to signal her sexual status when standing upright. Like modern woman (Masters and Johnson, 1966) and many other female mammals and primates such as the chimpanzee (Ford and Beach, 1951; Goodall, 1986, 1990, 1996, 2010), the human female likely retained the capacity to enjoy multiple sex partners (and to experience multiple orgasms) one after another, and to exercise sexual choice. This would have led to the selective breeding of intelligent, less violent males, sporting a big penis (Joseph 2000a,b, 2011).
It has also been estimated (Joseph 2000a,b, 2011) that the breasts and buttocks of the human female became permanently enlarged during this evolutionary time period, serving to continually signal her new sexual status and availability. Indeed, these same sexual signals are employed by other species when in estrus. In non-human primates, the vaginal area is directed in a posterior direction, and puffs out and becomes quite large and red or pink when she enters estrus. Thus, a red or pink, swollen derriere signals sexual receptivity and advertises the female's sexual availability. Some female primates, such as the gelada baboon, also advertise their sexual status via sexual swellings of the chest nodules which flush red (Fedigan, 1992; Jolly, 1972). These nodules form a necklace-like pattern which mimics the pattern of her rump--an enlarged derriere being a common sexual advertisement among many species.
However, in contrast to all other primates, the human female is the only female with permanently enlarged breasts and buttocks, and the only female who is in estrus 365 days a year and capable of sexual intercourse at all times. Non-human primates are fully sexually receptive only during circumscribe time periods, which coincides with enlargements of the breasts and the posterior directed vagina.
When female chimps and other apes and monkeys enter estrus, their dorsally oriented genitals/vaginal lips turn pink or a bright crimson and balloons outward, and in fact becomes so huge and distended they have difficulty sitting down. Moreover, estrus chimps and other primates, including baboons and the gorilla, go to great lengths to focus male attention on her buttocks, which she may flaunt and display by swaying them "enticingly" (Fedigan, 1992; Ford and Beach, 1951; Goodall, 1986, 1990; MacKinnon, 1979; Nadler, 1976; Wickler, 1973). As detailed by Joseph (2000a,b, 2011) the female genitalia and a derriere that is swollen or emphasized are obvious sexual signals which are employed to solicit male sexual attention. Likewise, modern human females accentuate and call attention to the derriere by wearing tight skirts and high heels which emphasizes the buttocks by puffing it out. When attired in this fashion she is assuming a sexually receptive posture and continually advertising her sexual availability.
The development of full time sexual receptivity and accompanying changes in the breasts and buttocks may also explain why H. erectus (or archaic H. sapiens) began utilizing various earth pigments (ochre), possibly for artistic or cosmetic purposes, or to emphasize female sexual availability. In one site, lumps of red ochre, many pointed like pencils, were found (Rightmire, 1990); and redness of the female primate genitalia is an obvious signal of sexual receptivity. This explains why modern women apply red lipstick, rouge, and other colorful cosmetics to their face and lips. The invention of cosmetics allowed women to purposefully mimic, mirror, and exaggerate the signs of estrus in other primates. However, this likely took place hundreds of thousands of years before the evolution of language (Joseph 2000a,b).
THE HOME BASE & LONG TERM ATTACHMENTS
These great changes in sexual status, the development of secondary sexual attributes and the invention of cosmetics to mirror the signs of estrus in other primates, most likely occurred during the rein of H. erectus, and contributed to the development of long term male-female mating relationships as is suggested by the creation of the home base and semi-permanent shelters where adult males and females and their young would reside (Joseph 2000a,b, 2011). That is, when female became full receptive, and could advertise this fact physically, dominant males were motivated to form long-term attachments. Normally, among most non-human mammals, males are interested in females only when they are capable of sexual intercourse, and lose interest when the estrus period wanes and disappears. Moreover, among many species of primate, dominant males may attempt to possess high status estrus females and prevent other males from mating with her, and will accomplish this by dragging her away to an isolated area which becomes a temporary home base.
For example, among chimpanzees, its not uncommon for a dominant male to threaten and physically force a high status estrus females to accompany him away from the troop, and to establish a home base where he provides her with an inordinate amount of attention; that is, until she ceases to be sexually receptive at which point he loses interest and returns to the troop (Goodall, 1986, 1990). Therefore, when the human female became continually sexually receptive, and continually advertised this fact, this may have motivated some human males to respond in a similar fashion and to form a long term mating relationship and to establish a personal home base, as is common among modern humans. Although a few other species mate for life, and/or limit their seasonal breeding to one mate, with the exception of gibbons, and some New World monkeys, this is not the case with other primates or most mammals, and is true in less than 1% of birds (Kleiman, 1977; Stacey, 1982; Wickler, 1973).
Hence, the emergence of full time sexual receptivity (and associated physical changes) likely contributed to the formation of the home base, long term mating relationships (or at least serial monogamy), and perhaps even the first "families" (Joseph 1993, 2000a,b, 2011). Thus, the emergence of sexual self-consciousness coincided with the invention of cosmetics, the evolution of breasts, buttocks and the big brain, and preceded the evolution of modern human consciousness and those tissues of the mind which produce and comprehend language and verbal-thoughts (Joseph 2011b).
CHIMPANZEES & AUSTRALOPITHECUS
The human female shares numerous sexual, cognitive, emotional, and behavioral characteristics with other species, the chimpanzee in particular (Bygott,1979; de Waal, 1989; Goodall, 1986, 1990; Itani & Suzuki, 2007; McGrew, 2011; Tanner, 2011, 1987; Stanford, 1998).
For example, although there is considerable variation, the sexual behavior of the female human (Gold & Burt, 1978; Matteo & Rissman, 1984; Udry & Morris, 1968, 1970; Wolfe, 1991) and the female chimpanzee (Fedigan, 1992; Michael, 1972) increases at the time of ovulation, that is, at midcycle; though in humans there is a second peak just before and after menstruation (Fisher, 1973).
Moreover, human and group living (multi-male/female) hominoid females are capable of experiencing multiple orgasms (e.g., Allen, and Lemmon, 2011; Burton, 1971; Chevalier-Skolnikoff, 1974; Goldfoot et al., 2010; Masters & Johnson, 1966; Michael et al., 1974) and commonly (though there are exceptions) have sex with multiple partners. Multiple sex partners ensures she will become pregnant, and multiple orgasms reward her with increasing pleasure as she sexually dallies with male after male.
These and other female chimpanzee/human commonalties appear to be due to a common genetic and evolutionary heritage and the sexual differentiation of the limbic system. It is this ancestral hominoid heritage and limbic system commonalties which explains not only her capacity for multiple orgasms, but the fact that the human female's "monthly rhythm of ovogenesis, ovulation, estrogen and progesterone secretion, uterine stimulation, and menstrual bleeding follows the basic primate pattern" (Beach, 1974, p. 356). Because they share common ancestors and a limbic system organized in a "female" pattern, female chimps and humans are sexually similar.
The human female sometimes behaves like, and biologically, in many respects is similar to a female chimpanzee in heat. This is not only because we share a common ancestor, but because our most distant "human" ancestors were essentially apes. Indeed, given these primate origins and commonalties, the initial evolution of the human females' unique sexuality is therefore best understood from a pongid perspective, beginning with Australopithecus, and later, H. habilis, whose social life was probably more ape-like than human-like.
The common ancestors for chimps and humans diverged 5 million year ago and the chimpanzee brain (395-410 cc) is just slightly smaller than the average Australopithecus brain (375-440 cc). Male chimps are also about 20% larger than females. Given these and the other genetic and sexual similarities mentioned above, it can be assumed that social-sexual and consort relations between ancestral males and females were more like that of chimpanzees.

Australopithecus vs Ape Skull
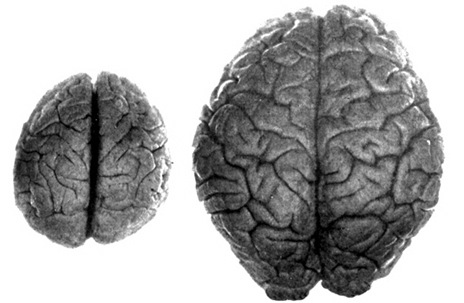
Australopithecus vs Human Brain
EVOLUTION OF FEMALE PELVIS AND THE "BIG BRAIN"
It was during the latter stages of H. erectus evolution that the brain became significantly enlarged. The brain in fact doubled in size with the transition from Australopithecus (440 cm3) to H. erectus (937 cm3), approaching within 15% of present-day humans (Conroy 1998; Potts, 1996; Rightmire, 1990; Tobias, 1971). However, with the advent of the big brain, human sex differences, including those related to full time female sexuality receptivity appear to have become even more pronounced.

Because a bigger brain comes in a bigger head, this required a larger birth canal and an increase in the sexual physical differentiation in the size and width of the H. erectus (and modern) female pelvis so as to accomodate the birth of a big brained baby (Joseph 1996, 2000a,b, 2011). Specifically, with the evolution of a bigger brain and with the transition from Australopithicus to H. habilis to H. erectus, the pelvic opening became longer and more round and ovoid as it expanded from front to back (Day, 1982, Lovejoy, 1988; Sigmon, 1982); changes with made birth more difficult as now the infant's head had to rotate as it passes down the birth canal in order to emerge from the pelvic opening.
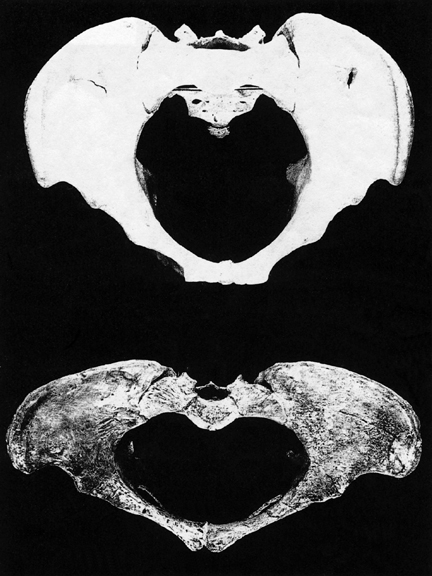
As with modern woman, this adaptation likely forced the female erectus' upper legs wider apart and her knees closer together, thus altering her gait and balance, causing her to wiggle her derriere when walking. Presumably, this alteration, coupled with the evolution of new muscles to aid in the upright stance, accentuated and drew attention to the female derriere and her sexual availability (Joseph, 1993; 2000a,b, 2011); a sexual-social signal accentuated in modern women through high heels and tight clothes, and in the last century, via the bustle, hoop skirts and in previous centuries dresses designed to grossly exaggerate the width of the hips.
These same sexual signals are characteristic of other primates (Ford and Beach, 1951; Wickler, 1973). When female chimps and other apes and monkeys enter estrus, their dorsally oriented genitals/vaginal lips turn pink or a bright crimson and balloons outward, and in fact becomes so huge and distended they have difficulty sitting down. Moreover, estrus chimps and other primates, including baboons and the gorilla, go to great lengths to focus male attention on her buttocks, which she may flaunt and display by swaying them "enticingly" (Joseph 2000a, 2011). The female genitalia and a derriere that is swollen or emphasized are obvious sexual signals which are employed to solicit male sexual attention. Likewise, modern human females accentuate and call attention to the derriere by wearing tight skirts and high heels which emphasizes the buttocks by puffing it out. When attired in this fashion she is assuming a sexually receptive posture and continually advertising her sexual availability.
THE FEMALE PELVIS AND THE DIVISION OF LABOR
The transformation of the human female hips and pelvis, however, also limited her ability to run and maneuver about in space, at least, as compared to most males (Joseph 2000a,b). This is because, be it H. erectus or modern H. sapiens sapiens "these changes are less pronounced in the... male pelvis" (Lovejoy, 1988, p. 123). "The size of the canal in men is controlled mainly by locomotor, not reproductive factors" (Campbell, 1985, p. 152).
Moreover, her pelvis became more fragile and subject to fracture if stressed by continous hard running and jumping. This condition plagues even female soldiers who suffer an unusually high incidence of pelvic and leg fractures although their training and duties are not as strenuous or arduous as males (2005 U.S. Marines Report, Paris Island). The female pelvis is in fact less massive, lighter and thus weaker than the male pelvis. Moreover, as the human femoral neck expanded and became longer so as to accommodate these changes in the pelvic girdle, it became more porous and thus weaker (Lovejoy, 1988) as well as subject to stress related injury due, in part, to the greater transverse and sagittal diameter of the female pelvic inlet.
As summarized by Day (1996, p. 5), "The female bony pelvis is a compromise between the needs of childbirth and those of locomotion with heavy selection pressure on the need for successful reproduction. The male pelvis, with no reproductive function to parallel that of the female, evolved for efficiency in bipedal locomotion." This helps "explain the differences in peak achievements between men and women in running and jumping."
Hence, be it modern woman or female H. erectus, in accomodating the birth of big brained babies, these physical changes not only advertised her sexual status, but likely interfered with her ability to endure and keep up with males on their long hunting sojourns; a function also of her reduced upper body strength and the prolonged dependency of her big brained babies (Joseph, 1993, 2000a,b, 2011). As such, with the evolution of H. erectus (and their descendants), males and females became increasingly specialized to perform certain tasks, e.g. hunting versus gathering--a division of labor characteristic of all hunting and gathering groups (Dahlberg, 2011; Hiatt, 1970; Martin and Voorhies, 1975; Murdock and Provost, 1973; Zilman, 2011) including chimpanzees (Goodall, 1990, 1996, 2010).
NATURAL BORN KILLERS: MALES ENJOY HUNTING & KILLING
Female primates, including chimps (whose activated DNA is 98.6% identical to human DNA), sate their hunger, and that of their infants, through systematic gathering of foodstuffs and insects, most of which is shared with their young (Goodall, 1990, 2010). Although male primates gather, they also hunt and become exceedingly excited when killing not only other animals (Hamburg, 1971; Harding and Strum, 1978), but other primates including former members of their troop who they may ambush and beat to death (Goodall, 1986, 1990). Be it other animals or a chimp from a neighboring band, when chimpanzees attack and wage war, and beat and kill other chimpanzees, the entire troop will gather and excitedly beg for a tiny morsel of the bloody flesh of victims as it is torn away by the victor (Goodall, 1986).


As is well established, be it human, non-human primate, or mammal, males tend to be violent (Elia, 1988; Fedigan, 1992; Goodall, 1986, 1990; Hamburg, 1971; Johnson, 1972; Lorenz, 1966; Manning, 1972; Mitchell, 1979; Moyer, 1974), and the male proclivity to hunt appears to be direct extension of these proclivities (Joseph, 1993, 2000a,b). Indeed, over 80% of all violent crimes and murders are committed by human males (Uniformed Crime Reports, 1990-1996, 2000-2006). Likewise, male chimpanzees initiate attack five times as frequently as females and are responsible for 90% of all aggressive encounters (Bygott, 1979; Van Lawick-Goodall, 1968).

Sex differences in violence and belligerence are apparent as early as the first two months of life even in chimps and other primates (Ransom and Rowell, 1972; Mitchell, 1979). Likewise, following the divergence between common ancestors, the human (male) tendency toward violence was probably also a characteristics of Australopithecus who not only hunted small animals, but possibly each other. According to Dart (1949), Australopithecines "were confirmed killers; carnivorous creatures that seized others by violence, battered them to death, tore apart their broken bodies, dismembered them limb from limb, and slaking their ravenous thirst with the hot blood of the pitiful victims and greedily devouring their writhing flesh." Although Dart's conclusions have been challenged, as noted above, male chimps not uncommonly engage in violent, murderous and even cannibalistic interactions with other troop member (Goodall, 1986, 1990).
Like modern humans, gangs of male chimpanzees will also engage in surprise attacks on neighboring colonies, beating and killing the old, infirm, and young alike, including former friends; even drinking their blood. That Australopithecus were killers and hunters of meat, thus, should not be surprising. In fact, from an examination of Australopithene teeth it is apparent that these were meat eaters whose diet also consisted of vegetable matter (Sponheimer & Lee-thorp, 1999)--as is the case of modern hunting gathering groups (Dahlberg, 2011; Lee and DeVore, 1968; Martin and Voorhies, 1975; Murdock and Provost, 1973; Zilman, 2011) and chimpanzees.
Specifically, from an analysis of the tooth enamel of Australopithecus africanus, it was determined that these individuals ate food rich in carbon 13--that is high protein food (Sponheimer & Lee-thorp, 1999). As these teeth were lacking the telltale scratches associated with vegetable matter, it can thus be concluded they were eating meat. Moreover, the remains of A. garhi were found in association with antelope leg bones which show the signs of cutting and pounding by a shark and blunt rock (White et al., 1999). Hence, it is fairly obvious that these were meat eaters and probably killers of animals and each other, as is the case with chimpanzees.
Thus sex differences in aggression and trends in the division of labor had probably been established with the evolution of hominoids (as is suggested by modern day chimps), and likely continued to develop with the emergence of Australopithecus. These same trends, however, were likely exaggerated with the evolution of the big brained H. erectus.
A bigger, more complex brain confers upon the bearer increased cognitive and intellectual capabilities, and the male H. erectus employed that greater intelligence in the pursuit of bigger game, including deer, bisons, horses and even bear and elephant (Potts, 1996; Rightmire, 1990). His killing technique, however, remained rather primitive. He would either surround his prey and stone them to death, or would use fire to frighten and stampede all manner of animals over cliffs or into swamps, typically killing more animals than could possibly be eaten (Potts, 1996; Rightmire, 1990).
The Silent Hunter Hunting does not require language (Joseph, 1993, 2000a,b). Rather, successful hunting requires prolonged silence, excellent visual-spatial and gross motor skills, and the capacity to endure long treks in the pursuit of prey. These are abilities at which males excel, including modern H. sapiens sapiens (Joseph, 1988a, 2000a,b). As detailed below, it is not until the emergence of modern (Upper Paleolithic) H. sapiens sapiens, around 35,000 to 50,000 years ago, that "modern language evolved.
Females Love Babies Just as primate males hunt whereas most primate females are not so inclined, it is unlikely that the female H. erectus, burdened by pregnancy, crying children, and her fragile pelvis and awkward gait, would have behaved otherwise. Rather, whereas most primate males, including male humans tend to have little interest in infants or caring for the young (Belsky et al., 1984; Clarke-Stewart, 1978; Frodi et al., 1982; Joseph 2000b) it is likely that the female H. erectus, like other female primates, would have pursued her own inclinations including the desire to bear babies and nurture the young.
In fact, be it chimpanzee, baboon, rhesus macaque, or human, females demonstrate an extraordinary interest in babies and will engage in play-mothering during even the earliest phases of their own childhood (Blakemore, 1990; Devore, 1964; Elia, 1988; Fedigan, 1992; Goodall, 1971, 1990).
Female humans (Berman, 1983; Blakemore, 1990) and non-human (group living) female primates will eagerly cuddle, groom, and hold babies that are not their own (Jolly, 1972; Kummer, 1971, Strum, 1987; Goodall, 1971). Although there are exceptions such as the pigtail macaque, in general, female humans, apes, or monkeys with babies, become the center of female-attention (Elia, 1988; Fedigan, 1992; Jolly, 1972, Mitchell, 1979, Strum, 1987).
Males are Indifferent Toward Babies By contrast, human men and boys and non-human male primates have little interest in babies or young children and generally provide little or no nurturant care even for their own offspring though they may form alliances with siblings (Fedigan, 1992; Joseph 2000a; Kummer, 1968, 1971; Mitchell, 1968, 1979, Goodall, 1971; Gordon and Draper, 1982; Rossi, 1985; Rowell et al., 1968), one of the few exceptions being owl monkeys, and to a lesser extent, baboons. Human males and fathers rarely behave in any manner that approximates normal female maternal behavior (Belsky et al., 1984; Clarke-Stewart, 1978; Frodi et al., 1982). As noted, males tend to be aggressive and violent and the desire to nurture infants may not be compatible with these tendencies. Indeed, males are responsible for over 70% of all murders involving infants and children (U.S. Justice Dept. 1996-1997, 2006-2007). Likewise, among many other primates (gorillas, Barbary apes, and rhesus, howler, and red-tailed monkeys) males stereotypically kill infants not their own (Fedigan, 1992; Hrdy, 1979).
THE INFANT-MATERNAL ROOTS OF EMOTIONAL LANGUAGE: BABIES & MOTHERS TALK TO EACH OTHER
Considerable vocalizing occurs between mothers and infants. The infants of many species will often sing along or produce sounds in accompaniment to those produced by their mothers (Barnett, 1995; Bayart et al., 1990; Jurgens, 1990; Wiener et al., 1990). These interactions reinforce and promote mutual vocalization, attachment, and contribute to survival. Hence, the first forms of complex social-emotional communication may have been produced in a maternal context (Joseph, 1992, 1993, 1996, 1999).
Because the big brained infant H. erectus likely required long term care, it can be assumed that there was an impetus for female H. erectus to vocalize with her young for years at a time. Indeed, a big brain requires more time to grow and mature, which in turn results in prolonged immaturity and helplessness. In consequence, the female tendency to vocalize more than males may have been given additional impetus well over 500,000 years ago.
Among most social animals and gathering groups, the production of sound is very important in regard to infant care. If an infant becomes lost, separated, or in danger, this is best conveyed via a cry of distress and fear; cries which would cause a mother to come running to the rescue. Conversely, vocalizations produced by the mother and her co-gatherers enable an infant (or a lost gatherer) to orient and find its way back if perchance it got lost or separated. In this regard, the tendency to vocalize may have ensured for breeding success.
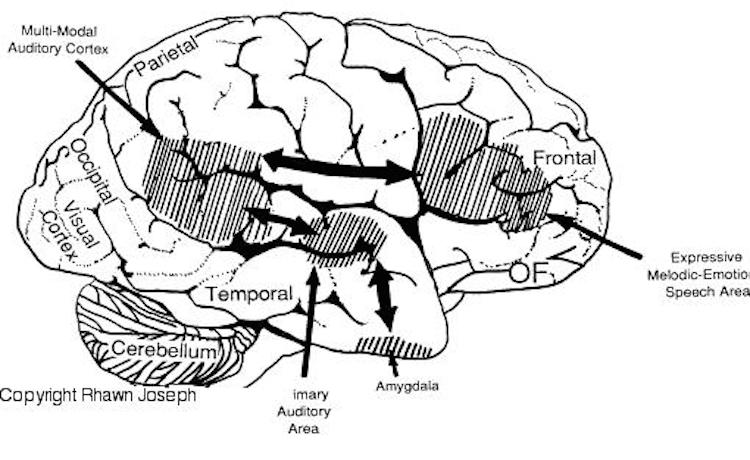
Among animals, and present day human mothers, much of this initial mutual sound production is emotional and prosodic (Cooper and Aslin, 1990; Fernald, 1991, 1992; Fernald et al., 1989; Jurgens, 1990) and constitutes what has been referred to as "limbic language" (Joseph 1982, 2000). Emotional sound production is mediated by the amygdala, cingulate gyrus, and other limbic and subcortical nuclei, but is also hierarchically represented in humans and produced and perceived by the right frontal and right temporal-parietal area. These emotional-language areas appear to be more extensively developed in human females (Joseph 1993, 1996, 2000b). By contrast, the denotative, and vocabulary-rich grammatical components of modern human speech are mediated by the left frontal and temporal-parietal area; i.e. Broca's and Wernicke's speech areas and the inferior parietal lobule.
Because infants are emotionally oriented and have little or no understanding of non-emotional speech, these mutual mother-infant vocalizations usually consists of exaggerated emotional prosody (Cooper and Aslin, 1990; Fernald, 1991, 1992, 1993; Fernald et al., 1989). Indeed, human infants prefer listening to and are more responsive to these exaggerated emotional vocalizations, particularly when produced by a female (Cooper and Aslin, 1990). Moreover, human (and non-human) females demonstrate superiorities in this regard, and not only produce more mutual social-emotional vocalizations than males (Brody, 1985; Burton and Levy, 1989; Gilbert, 1969; Glass, 1992; Tannen, 1990) and more words in five seconds of conversation, but tend to employ 5-6 different prosodic variations and utilize the higher registers when conversing (Joseph, 1993). They're also more likely to employ glissando or sliding effects between stressed syllables (Brend, 1975; Coleman, 1971; Edelsky, 1979). Men tend to be more monotone, employing 2-3 variations on average, most of which hovers around the lower registers (Brend, 1975; Coleman, 1971; Edelsky, 2009). Even when trying to emphasize a point males are less likely to employ melodic extremes but instead tend to speak louder.
FEMALE & BILATERAL EMOTIONS
Although influenced by sex differences in the oral-laryngeal structures, these differential vocalizing abilities are also reflected in the greater capacity of the female brain (and right hemisphere) to express and perceive emotional vocalizations (Burton and Levy, 1989; Hall, 1998; Soloman and Ali, 1972). This superior sensitivity includes the ability to understand, perceive, and express empathy and social-emotional nuances (Burton and Levy, 1989; Brody, 1985; Buck, 1984; Eisenberg et al., 1989; Fuchs and Thelan, 1988; Harackiewicz, 1982; Lewis, 1983) and a greater willingness to express emotional issues and discuss personal problems (Gilligan, 1982; Joseph 1993, 1996, Lutz, 2010; Walker et al., 1997; Lombardo and Levine, 2011).
In fact, from childhood to adulthood, females appear to be much more emotionally expressive than males (Brody, 1985; Burton and Levy, 1989; Gilbert, 1999; Tannen, 2000), who in contrast have difficulty discussing personal difficulties or expressing their emotions other than through anger, happiness, and sexual arousal (Balswick, 1982, 1988; O'Neil, 1982; Sattel, 1999; Tannen, 2000).
FEMALE H. ERECTUS & LANGUAGE
Like chimpanzees and present-day human females, female H. erectus probably engaged in mutual mother-infant vocalizations, and probably engaged in more mutual social-emotional vocalization not just with her children but with each other. However, as based on an analysis of H. erectus' tool technology, and the fact that it was somewhat crude and lacking in temporal sequential sophistication (though certainly a lot of time was put into their construction), it can be assumed that the brain of H. erectus was not capable of thinking in terms of complex temporal sequences. Hence, it can be assumed that H. erectus did not speak in the temporal sequential and grammatical manner characteristic of present-day humans. Nor is it likely H, erectus uttered complex words.
Rather, H. erectus probably tended to use gesture, body language, facial expression, as well as grunts, groans, mimicry, and the production of emotional sounds in order to convey needs, fears, feelings and desires. On the other hand, as gathering and the construction of domestic-tools was probably an ongoing female H. erectus activity (see below), these activities likely eventually gave rise to improvements in bilateral, temporal and sequential fine motor skills; thus providing what would become the neurological foundation for the development of complex grammatical, vocabulary-rich, and emotional human language.
HUNTING VS DOMESTIC TOOLS
Whereas the big brained male H. erectus refined his hunting techniques and became a hunter of bigger game, females refined their gathering and food preparation techniques as is suggested by the creation of crude tools such as choppers, scrapers, cleavers with a stright cutting edge on one side (Day, 1996; Rightmire, 1990), and probably digging sticks, as is characteristic of female chimpanzees (Goodall, 1990; McGrew and Marchant, 1992). That these tools were fashioned by a female hand can be deduced by their domestic use. Among hunting and gathering groups it is females and not the males who make and use tools (Joseph 1992, 1993, 2000a,b; Niethhammer, 1977), the only exception being hunting implements and weapons of war which females are not allowed to touch (Tabet, 1982). Similarly, females chimps use food/gathering-related tools much more frequently than males (Goodall, 1986; McGrew and Marchant, 1992) who in turn are more likely to use sticks and rocks to threaten other males (Bygott, 1979; Goodall, 1990; Van Lawick-Goodall, 1968 ). As with other male primates, and given his penchant for big game hunting, it was probably the male H. erectus who created the first hand ax; a killing tool which made its appearance about 650,000 years ago (Potts, 1996; Rightmire, 1990).
As tools for "gathering" versus "hunting" were fashioned throughout Africa, Europe, Asia and the Middle East throughout the Paleolithic, it can be assumed, with the exception of the European Neanderthals, that this division of labor prevailed throughout the Middle Paleolithic, well after H. erectus was replaced by archaic H. sapiens (Joseph 1993, 1996, 2000a,b). For over 500,000 years amles have hunted and killed whereas females cared for the young, maintained the home base, and were responsible for the gathering of food and related "domestic" activities. However, this is not to imply that females never killed small animals, or never collaborated or assisted in the hunt, as that is not the case with humans or chimpanzees.
By the close the Middle Paleolithic (around 30,000 B.P.), and the emergence of "modern" Upper Paleolithic H. sapiens sapiens, such as the Cro-Magnon, hunting had become the center of religious and artistic life. Nevertheless, 60-80% of the Cro-Magon diet consisted of fruits, nuts, grains, honey, roots and vegetables (Clark, 1952; Prideaux, 1973), which were probably gathered by females (Joseph 1992, 1993, 2000a,b). Among the hunting and gathering societies in existence during the last few centurties, women have been and are the gatherers and main providers of food whereas spoils from the hunt account for only about 35% of the diet (Dahlberg, 2011; Lee and DeVore, 1968; Martin and Voorhies, 1975; Murdock and Provost, 1973; Zilman, 2011).
FEMALE TOOL-MAKERS, TEMPORAL SEQUENCING AND LANGUAGE
In grubbing for roots and bulbs the Upper Paleolithic female-gatherer probably used a digging stick which she periodically sharpened by using stone flakes. These gatherers also fashioned hammerstones for cracking nuts and grinding the produce she collected. As in recent hunting gathering groups (Lee, 1974; Murdock and Provost, 1973), her duties would include the preparation of any meats she scavenged or which the men brought home from their hunting sojourns. In addition to food preparation, clothes were sewn and fashioned out of hides (Clark, 1952, 2007; Prideaux, 1973), and these too are tasks associated with women (Gusinde, 1961; Lee, 1974; Neithhammer, 1977), including, presumably, the Cro-Magnon females of the Upper Paleolithic (Joseph 1992, 1993, 1996, 2000ab).
Thus the duties of the Upper Paleolithic female were much more multi-faceted and complex than her predecessors, and included cleaning hides via a scraper, drying and curing the skin over the smoke of a fire, and then using a knife or cutter to make the general desired shape. The Upper Paleolithic female was also employing a punch to make holes in these hides, through which leather straps or a vine could be passed so as to create a garment that could keep out the cold (Clark, 1952, 2007; Prideaux, 1973). They were also weaving and using a needle to sew garments together; "domestic" tasks which are almost exclusively associated with "women's" work (Murdock and Provost, 1973; Neithhammer, 1977).
The necessary skills required for tool construction and the efficient gathering of vegetables, fruits, seeds, berries and the digging of roots, include rapid, temporal-sequential fine motor maneuvers with the arms, hands, and particularly the fingers (Hamrick et al., 1998; Marzek, 1997; Susman, 1994). Similarly, most tools are made in a step-wise, temporal sequential manner, with specific fine-motor movements, and considerable precision; abilities which enable the tool maker to construct the same implement over and over again. This also requires that the tool maker be in possession of a brain that can control the hand and which can use foresight and planning in order to carry out the steps involved in the implement's manufacture. In this regard, although males were also fashioners of tools (i.e. those used for killing), it is noteworthy that females tend to excel at fine motor activities, such as those involving rapid, repetitive temporal sequencing (Broverman et al., 1968; Hampson and Kimura, 1992; Junaid and Fellowes 2006; Larson et al. 2007; Thomas and French 1985).
These "domestic" activities, coupled with her innate tendency to engage in mutual vocalizations, and to vocalize with her young and fellow gatherers, likely coincided with, and stimulated the development of the temporal sequential neurological foundations of what would become a female superiority in the evolution of grammatical, vocabulary-rich human language. However, as the tools made by H. erectus and archaic (Middle Paleolithic) H. sapiens remained relatively crude, whereas a creative explosion in tool-making technology typifies the onset of the Upper Paeleolithic (Clark, 1952, 2007; Joseph 1992, 1993, 1996, 2000a,b; Mellars, 1989, 2010; Prideaux, 1973), it appears that the human brain did not become fully adapted for perceiving and expressing temporal sequential, grammatically complex, vocabulary-rich language, until the emergence of the Upper Paleolithic human female. As will be detailed below, the emergence of these linguistic capabilities were promoted by and thus coincided with advances in tool technology and associated fine motor skills--activities which gave rise to the functional evolution of Broca's speech area and the angular gyrus of the inferior parietal lobe.
SEX DIFFERENCES IN LANGUAGE
Over the course of the last 500,000 years, women have been engaging in group and domestic activities including child rearing, that promoted the evolution of the neural substrates which subserve the development of human speech, including bilateral representation of language.
Language is both emotional and descriptive, grammatical and melodic, consisting of word units and prosody, such that just as both halves of the brain interact to produce music (Joseph, 1982, 1988), both halves become activated when engaged in language related activities, the left providing the grammatical word units, the right supplying the emotional melody. Hence, as demonstrated through functional imaging, the right (as well as the left) hemisphere becomes activated (Bookheimer, et al., 1995; Bottini et al., 1994; Peterson, et al. 1988; Price, et al., 1996; Shaywitz, et al., 1995). However, the female right (and left) hemisphere appears to be superiorally endowed with these and other aspects of language (Joseph, 1982, 1993, 1996; Shaywitz et al., 1995). Females demonstrate bilateral cerebral activation when engaged in certain language tasks as demonstrated by functional imaging (Shaywitz, et al., 1995).
Over 50 years of research has clearly demonstrated that human females display superior language, articulation, word knowledge, syntactic, and related linguistic skills as compared to males (Bayley, 1968; Broverman et al., 1968; Harris, 1978; Koenigsknecht and Friedman, 1976; Hampson and Kimura, 1992; Harshman et al., 1983; Hyde and Lynn, 1988; Kimura, 1993; Levy and Heller, 1992; Lezak, 1983; McGlone, 2010; McGuiness, 1976; Moore, 2007). In contrast to males, human females vocalize more, engage in more social speech, display superior linguistic skills, and excel over males on word fluency tests, for example, naming as many words containing a certain letter, or words belonging to a certain category. Females also vocalize more as infants, speak their first words and develop larger vocabularies at an earlier age. Their speech as children is easier to understand, they improve their articulation and grammatical skills at a faster rate, and the length and complexity of their sentences is greater than males (Bayley, 1968; Harris, 1978; Hyde and Lynn, 1988; Koenigsknecht and Friedman, 1976; Levy and Heller, 1992; Lezak, 1983; McGlone, 2010; McGuiness, 1976; Moore, 2007).
Moreover, women gravitate toward and are overwhelmingly over represented in professions requiring conversational language, such as telephone operators or secretary--over 90% of those working in these fields are women (U.S. Department of Labor, 1997, 2007).
Whereas women and girls display clear language superiorities, or a greater tendency to vocalize and speak, males suffer from more language related disturbances such as stuttering (Corballis and Beale, 1983; Lewis and Hoover, 1983). Moreover, males lose language-related capabilities as they become aged, are more likely to become aphasic following stroke, and do not recover language as quickly or as fully as females (Joseph 1996).
In addition, girls learn how to read more quickly and more proficiently than males who are more likely to suffer from reading difficulties including dyslexia (Corballis and Beale, 1983). Females not only demonstrate superior reading comprehension and writing and spelling skills (Lewis and Hoover, 1983), but in 1997 the U.S. Department of Education (USDE) reported that the writing skills of a 9 year old girl in fourth grade is equal to that of a 13 year old boy in 8th grade, and in 2007 the USDE reported that "at all age levels females continue to outscore males in reading proficiency... and females of all ages have outscored males in writing." Rreading and writing are directly associated with the functional integrity of the IPL/angular gyrus.
Presumably, females demonstrate superior language skills because their brains are better adapted for producing temporal-sequential and social-emotional vocalizations (via the maternal cingulate gyrus), and because, as gatherers and tool makers they were not under the same constraints as the silent male hunters. Males were required to maintain long periods of relative silence -at least while hunting. Their brains adapted accordingly.
GATHERING & WHY WOMEN LIKE TO TALK, SHOP, AND SHARE THEIR FEELINGS
Over the course of human evolution, and in contrast to the silent, non-communicative (or threatening) males, female mothers and female gatherers were able to freely chatter with their babies or amongst themselves. Indeed, gathering not only fostered the development of language, but like hunting for men, also served as a social activity that enabled women to interact in a socially intimate manner.

Our ancient female ancestors probably gathered in large groups of 7 or more individuals, as the typical size of a band is about 25 individuals on average. Some women were pregnant or probably carried infants which might be set on the ground here and there, or accompanied by young adolescents who would frolic about and play. Such gathering groups must have commonly been loud, noisy and very gay affairs filled with the talk of the women and the sounds of games and yells of the children. Hence, unlike the men who must remain quiet for long time periods in order to not scare off game, the women are free to chatter and talk to their hearts delight. Talking also served as a means of maintaining the location of the group so that if a gatherer or a child chanced to walk away, she (or her child) could always relocate the others by their hodgepodge of speech. However, it is also likely many women did not wonder far away from the group or home base as they might become prey.
Talking thus became part of the gathering glue and served the purpose of keeping the group together and thus of bonding the group as a collective (a function similar to that performed by the anterior "maternal" cingulate gyrus). The women talked about their children, their mates, each other, and were thus exposed and allowed to expose their own feelings and thoughts to those who valued talking as much as they. As will be detailed below, for women, to socialize and be together means to talk, and to talk is a very social bonding element for women even today (Joseph 1992, 2000b; Tanner 1992); a characteristic first forged by the evolution of the anterior "maternal" cingulate gyrus of the limbic system and the subsequent female tendency to vocalize with her young (Joseph 1996).
When considering our evolutionary heritage and the fact that women have spent a good 100,000 years or more in female dominated gathering groups, where socializing and freely expressing oneself about social and family matters were the norm, it should come as no surprise that modern day females continue to respond similarly. It is thus little wonder that some women also feel a compulsion to shop and derive considerable enjoyment making it into an all day affair. Likewise, some modern men, bravely armed with high powered weapons, also feel a compulsion to stalk and kill helpless animals, or even other humans. It is hard to escape hundreds of thousands of years of evolutionary pressure.
In this regard, it perhaps should not be surprising that after almost half a million years of holding his tongue and jockeying for status and position amongst his fellows, that modern day man continues to respond similarly and have the same concerns, albeit translated and modified to some degree; i.e. sports, politics, business.
However, it is also because woman has engaged in group gathering activities, accompanied by sisters, daughters, cousins, and friends, for over a million years, that "modern" westernized woman continues to feel an innate need to "gather." That is, she "shops" and derives considerable enjoyment from shopping. Over 90% of department store space is devoted to women, because of this innate female characteristic. Women may shop when they are depressed and lonely--as gathering was always a social activity-- and they may buy dozens of articles, and "cute little shoes" and other clothes that they may never wear or wear only once. She feels impelled to shop/gather. Indeed, a woman can spend hours "shopping" and might make it an all day affair. By contrast, a man will often go to the store, pick up what he needs and leaves. A woman gathers and gathers along side numerous other gathering/shopping women. Men hunt until they kill and then go home.
Women gather. Men hunt.
The nature of human consciousness has been shaped by our evolution history.
HUNTING, VISUAL-SPATIAL SKILLS, & WHY MEN DON'T SHARE THEIR FEELINGS
Searching for large game animals requires that the hunting band roam quietly over a huge expanse of varied terrain which may extend up to 500 or more miles. To be successful requires good directional sense, so that one could wonder and also find their way home without the aid of a street sign, good visual-spatial skills such as depth and distance perception, so that one can walk and run as well as anticipate the trajectory of movement of a running prey, without tripping and falling, an excellent capacity to recognize, comprehend, and mimic animal, environmental and non-speech sounds, and an increased capacity to communicate non-verbally via gesture, body language, and particularly via animals mimicry so that game would not be scared off by the sounds of speech and so that communication with other hunters could occur. These are all skills associated with the right hemisphere of a modern day human being.
Indeed, the mimicry of certain animal sounds could communicate a host of meanings such as where something might be located (as those animals whose sound he is mimicking may be found only in a certain location), that a particular beast (which elicits cries from the creature being mimicked) is approaching or nearby, or a particular action that the group should take (fight or flight or freeze), and so on. These are all capacities at which the right hemisphere of modern man and woman, excels.
However, in that big game hunting was an activity dominated by men, when one considers the skills involved to be a successful big game hunter it should be no surprise that after 300 or more thousand years of engaging in these activities, that males continue to demonstrate cognitive superiorities in regard to related abilities.
When males banded together for the purposes of the big game hunt, they had to walk silently for long time periods and their only form of communication was probably concerned only with the expression of facts and information about the hunting they were engaged in. If they were to begin talking about anything else such as their family, they possibly would have been shunned as their discussion would have been irrelevant, and would have scared off the game. A premium was thus placed on men who could hold their tongue.
Also decreasing the need for talk was the fact that as hunters stalk their prey they may walk at anywhere from 25-100 yards apart. Two or more men standing together is more likely to warn a prey of their presence, whereas men standing apart have the advantage of chasing a beast into the waiting arms of his comrade.
Finally, it is likely that once the hunt was complete and they returned to their base camp, that the men did not then begin to discuss their wives, children, feelings, or personal problems, but their own prowess as hunters, incidents related to the hunt, and whatever adventures they may have had that would enhance their status amongst their fellows.
VISUAL-SPATIAL SKILLS OF THE SILENT HUNTER
In contrast to females where language representation appears to be bilateral, language appears to be more greatly concentrated in the left half of the male brain (Bradshaw et al. 1977; Joseph, 1982, 1993, 1996; McGlone, 2010; Shaywitz et al., 1995). For example, among males only left hemisphere lesions result in these language deficits whereas women are more likely to suffer word finding difficulty and a reduction in vocabulary with right or left hemisphere lesions (Kimura and Harshman, 1984). In contrast to the female cerebrum, the right half of the male-brain appears to be more lateralized and functionally specialized to analyze (and sequence) visual-spatial relationships (Levy, 1972, 1974; McGee, 1979; McGlone, 2010; Witelson, 1985)--perceptual functions directly related to skill at hunting.
Whereas female gatherers are free to talk to one another and their children, hunting does not promote fine motor skills or linguistic development as the successful hunter must be silent and capable of throwing a spear and analyzing the spatial coordinates which separate yet link him to a potential prey. These male activities do not promote speech. Wolves and wild dogs spend a considerable part of each morning and evening tracking and hunting and there is no evidence of speech among these creatures. Speech confers few advantages to a group of human hunters, who must maintain long periods of silence so as to not scare off potential game.
Aspects of hunting, however, also put a premium on parietal-temporal lobe and right cerebral cognitive development. Tracking, aiming, throwing, geometric analysis of spatial relationships, as well as environmental sound analysis, are also directly related to the functional integrity of the right half of the brain (Guiard, 1983; Haaland and Harrington, 1990; Joseph, 1982, 1988a,b, 1996) and the parietal lobe in particular. The right parietal area is associated with the mediation of visual-spatial perceptual functions, especially in males, including providing the sensory feedback which would enable a hunter to aim and throw a spear; guidance which is transferred, via the corpus callosum to the left hemisphere somatomotor areas controlling the right upper extremity.

Hence, given 500,000 years of multi-generational male experience in hunting which usually required days or even weeks of wondering hundreds of miles from the home base, present-day males therefore demonstrate superior visual spatial skills including superior maze learning, tracking, aiming, and related non-verbal abilities, as compared to females (Broverman et al., 1968; Dawson et al., 1975; Harris, 1978; Joseph, 1993, 1996; Kimura, 1993; Levy and Heller, 1992). This includes a male superiority in the recall of geometric shapes, detecting figures that are hidden and embedded within a complex array, constructing 3-dimensional figures from 2-dimensional patterns, visually rotating and detecting the number of objects in a 3-dimensional array, and playing and winning at chess (which requires superior spatial abilities).
Males possess a superior geometric awareness and directional sense and geographic knowledge, are better at solving tactual and visual mazes or mentally manipulating spatial relations on paper, and are far superior to females in aiming, throwing, and tracking such as in coordinating one's movements in relationship to a moving target (Broverman et al., 1968; Harris, 1978; Kimura, 1993; Levy and Heller, 1992; Linn and Petersen, 1985; Porteus, 1965; Thomas et al., 1973). Only about 25% of females in general exceed the average performance of males on tests of such abilities (Harris, 1978; Joseph 2000a).
Moreover, some of these differences are present during childhood and include a male superiority in the recall and detection of geometric shapes, detecting figures that are hidden and embedded in an array of other stimuli, constructing 3-dimensional figures from 2-dimensional patterns, visually rotating or recognizing the number of objects in a 3-dimensional array, playing and winning at chess (which requires superior spatial abilities), directional sense and geographic knowledge, the solving of tactual and visual mazes, aiming and tracking such as in coordinating one's movements in relationship to a moving target, coordination in aiming and throwing, and comprehending geometrical concepts. Many of these abilities are directly related to skills that would enable an ancient hunter efficiently to track, throw a spear, and dispatch various prey without getting lost and while maintaining a keen awareness of all else occurring within the environment.
Various aspects of mathematical ability are directly related to visual-spatial functioning, such as geometry and the sequencing of space (Joseph 1988a; 1990, 1996). However, whereas the female brain has an advantage when it comes to sequencing language, the male brain is more adept at sequencing space. Hence, males are far more likely than females to be mathematical geniuses. Even when the sample consists of females who are scholastically gifted, males achieve higher scores, for example, on the math portion of the college entrance exam, and outperform females by a ratio of 13 to 1 (Benbow and Benbow, 1984).
Moreover, some of these differences are present during childhood and early adolescence and have been demonstrated in other species (Linn and Petersen, 1985; Joseph, 1979; Joseph and Gallagher, 2010; Joseph et al., 1978). For example, male rats consistently demonstrate superior visual-spatial and maze learning skills as compared to females (Joseph, 1979, Joseph and Gallagher, 2010; Joseph et al., 1978). If reared in an enriched environment, males continue to outperform females (Joseph, 1979; Joseph and Gallagher, 2010).
SEX DIFFERENCES IN COMMUNICATION
It has frequently been noted that many men tend to interact and compete with one another in terms of power, status, physical and intellectual dominance, wealth, and control. In contrast, it has been argued that women tend to speak and interact in a more cooperative manner, and to be more interested in issues related to the family, inter-dependence, and social intimacy (Croates, 1986; de Beauvoir, 1961; Joseph, 1985, 1992; Stutman, 1983; Tanner, 1990). Although as a rule, such notions do not apply to all men or women, there indeed does appear to be a gender linked difference in the manner in which many boys, men, women, and girls tend to interact and speak together; and these same sex differences appear to be biological in origin, directly related to the hunting vs gathering way of life which has characterized much of human evolution, and are also seen in other primates such as chimpanzees.
As with chimpanzees, many boys tend to play in groups where there is a recognized leader and hierarchical order of followers. The leader often wins this position based on physical strength and capability, risk taking, and his ability to control or bully others. Similarly, the hierarchy of followers is arranged along these same competitive lines. When playing boys often tend to engage in sometimes very physical, aggressive games, where wrestling, tripping, and pushing each other in fun is part of the activity (Brooks-Gunn & Matthews, 1979; Eder & Hallinan, 1978; Lever, 1976; McGrew, 1979; Savin-Williams, 2010). That this is rooted in biology and not early training, however, is also indicated by the fact that male chimpanzees and other social mammals behave in a similar fashion (de Waal, 1989; Eibl-Eibesfeldt, 1995; Goodall, 1986, 1990; Wickler, 1973).
Many girls tend to play in much smaller groups or in pairs; i.e. best friends (which is equivalent to mother-baby, sister-sister). Although hierarchies also tend to form, rather than based on physical competitiveness (although that is often a factor) it tends to be based on personality, articulatory skills, and physical attractiveness; e.g. who is the "nicest," prettiest, or the most fashionably dressed (Brooks-Gunn & Matthews, 1979; de Beauvoir, 1961; Eder & Hallinan, 1978; Ekert, 1990; Joseph, 1985). It is precisely because women are also competitive (see chapter 8), that female relationships with females who are not kin, tend not to be as long lasting, as compared to male-male friendships, the exception being female-female (e.g. mother-daughter, sister-sister) kin relationships, which, however, also tend to be quite rocky at times.
In contrast to boys and men who are more likely to become involved in physically aggressive team sports where there are clear winners and losers, many young girls are more likely to engage in cooperative activities that focus on friendship, intimacy, sharing, talking, imaginativeness, and being liked. And even among female athletes, the emphasis is often on the cohesiveness and importance of the team as a mutual cooperative, whereas males often seek to emphasize their own importance and personal status and accomplishments and to belittle their competitors.
Challenges and competition between girls are more likely to be subtle and indirect, whereas cooperativeness, at least overtly, is the glue which binds them together (Eder & Hallinan, 1978; Ekert, 1990). That this is biologically based is evident in that female chimpanzees behave in an identical fashion. As summarized by De Wall (1989, p. 137): "Adult male chimpanzees seem to live in a hierarchical world with replaceable coalitions partners and a single permanent goal: power. Adult females, in contrast, live in a horizontal world of social connections. Their coalitions are committed to particular individuals whose security is their goal...for them it is of paramount importance to keep good relationships with a small circle of family and friends."
Although both girls and boys often are concerned with being the best and tend to use force in order to get their way, boys are more likely to utilize actual physical violence; a trait rooted in biology, the sexual differentiation of the male brain, and the fact that males in general, almost regardless of species are more violent and murderous than females and in fact employ violence in order to gain access to females. By contrast, females tend to use words as weapons (the proverbial "woman's tongue") and rely on males to behave aggressively for them, and employ their sexuality to manipulate men or to compete with other women.
It is important to emphasize, however, that there are numerous exceptions, as many girls are quite physically competitive and aggressive, and many boys seek close social and emotional intimacy with their best friends as well. Probably most members of both sexes, in fact, fluctuate between these various different modes of interacting.
Nevertheless, these general patterns of interaction in turn tend to color the way in which men and women interact as adults as well, including the manner in which they view the actions and even the speech of others. That is, in very general terms, men and women often tend to be concerned with different aspects of the same experience since they sometimes have different priorities. In fact, even when males and females are engaging in ostensibly the same conversation, they focus on different aspects of the same shared information and emphasize features which are quite distinct (Croates, 1986; Emil & Hallinan 1978; Tannen, 1990).
Many males tend to employ speech and language as a means of imparting not only information, but as a manner of establishing status and superiority, which in turn may promote a man's ability to gain access not only to resources, but female sex partners. It is a common male stratagem to continually probe for weakness and to make continual attacks until thwarted or power is gained. However, such fights need not always be physical but based on threat and bluff and dominance displays that entail the clever or derogatory use of language. For example, human adolescent and adult males often utilize sarcasm, teasing, verbal ridicule, verbal directives, sexual jokes, or direct insults so as to establish superiority over a rival who may whither in response to the continual verbal onslaught. Indeed, sexual remarks, sexual "jokes" and teasing are frequent among adolescents as well as "professional" men, as it is a means of establishing sexual dominanc.
Among men, comments about one another can be quite graphic and often go well beyond innuendo and include remarks as to sexual inadequacy or potency, and may go so far as to challenge other males to serve as willing sexual orifices; e.g. "Suck my dick!" Insults and sexual comments are in fact often seemingly made in fun, and although ostensibly and overtly accepted as such, they usually belie attempts to achieve dominance over other men (Joseph, 1985, 1992b, 1993).
In fact, sexual threats, with direct exposure of an erect penis is often employed by primates in their dominance displays, whereas phallic displays are a "universal" expression among human societies (Eibl-Eibesfeldt, 1995; Wickler, 1973). Indeed, in part is because of the potentially threatening nature of the penis, that phallic aggression has crept into the vocabulary. When someone is really "pissed off" we know he is angry and we should watch out. If someone says "fuck you!" we know they are not inviting us to have sex. If someone has been "fucked", or "screwed" it is understood that they have been diminished or taken advantaged of in some manner.
Of course, males also use phallic and other forms of rough, derogatory and teasing comments in order to establish comraderie. For example, one male friend may say to another, "you still driving that piece of shit," and both will laugh. However, when a man teases, comments, or interacts with a woman on a similar level, even when it is extremely and considerably toned down, she is likely to become exceedingly offended, sometimes to the bewilderment of the man who sees his behavior as normal.
Females are much less likely to insult one another in fun, deride each other's supposed sexual shortcomings (at least while face to face) or demand in a jocular tone that other women serve them as sexual objects. Unlike some men, it is not seen as as a means of establishing rapport or dominance.
Moreover, when women engage in the seeking of status, or when they seek to put one another down (again, at least while face to face) it tends to be less confrontational, more subtle and less aggressive as ostensibly they appear more concerned with the social harmony and the establishment of mutual, friendly understanding and rapport (Hughes, 1988). They are more likely to try to smooth things over and be more artful and subtle rather than overtly or physically aggressive, even when jockeying for positions of dominance. Of course, like men, many women have no difficulty with cutting up a competitor.
Men and women thus tend to use language differently and in order to convey different messages. Although both use language in order to convey information, details and facts, many women tend to be more interested in discussing what they or others feel, what happened to them or their friends, who said what, and how various relationships are going. For many women, talk often serves as a means for maintaining intimacy and friendly interaction as well as for conveying details on business, finance, and politics and a variety of other subjects (Croates, 1986; Joseph, 1992b, Tannen, 1990). For many men, these personal details are inconsequential and of little importance.
Andy (yawning and checking his email): "So what happened?"
Ginger: "Turn off that computer so I can tell you. Anyway, she has lost so much weight. She must really be feeling good about herself, with all the money she has been bringing in. Even Sally was impressed, and you know how Sally feels about that."
Andy: "About what?"
Ginger: "Haven't you been listening to me?"
Andy: "Yeah, sure. Business! lunch! I'm listening. Go on."
Ginger: "Anyway, and, oh, did I tell you? Sally's little boy has been getting so big. She brought these pictures. He is just so cute. And Ruth..."
Andy: "I thought this was a business lunch."
Ginger: "It was. You should have seen the look on Ruth's face when she saw those pictures. She wants to have a baby so bad...."
Andy: "What has this got to do with your job? What did your boss have to say about that?"
Ginger: "Sally is my boss!"
Andy: "And she brings pictures of her kid to show you and her clients?"
Ginger: "What's wrong with that?"
Andy: "I don't see what that has to do with your job."
Ginger: "It has everything to do with my job. If you work with people you should be interested in what is going on in their life. People don't live and work in a vacuum. What goes on at home affects their work. Haven't you've learned that ?
Andy (yawning and clicking on Sports News): "Learned what?"
For many men, when language and talk is personal it is either viewed as irrelevant or power oriented. It often becomes personal when they feel their status or authority may be threatened.
Jake has come home and begins to complain to his wife Ruth about a problem at work. He is very irritated and upset.
Jake: "I don't know what's wrong with my boss. He hired this new girl and she is not working out!
Ruth: "What does she look like?"
Jake: "Very ugly. Anyway, she was supposed to type this stuff and she never even got started.
Ruth: "How old is she? Is she pretty?"
Jake: "What's that got to do with anything? She's not doing her job. She was supposed to type these papers for me and she didn't do it."
Ruth: "Well, she's new. Is she married?"
Jake: "Why are you asking me all this crap? Who cares what she looks like or how many times she's been married. She is incompetent."
Ruth: "Maybe if you were nicer to her..."
Jake: "Hey! Whose side are you on? She's the one who is screwing up. Not me."
Ruth: "I didn't say you were."
Jake: "Bull shit! I don't know why I even bother talking to you. All you do is put me down."
Ruth: "Well excuse me for being curious."
By contrast, those men who like to gossip about others and to share intimate and personal details are often shunned by other men as they are not interesting to talk to.
Women are more likely than men to use language as a means of maintaining social intimacy and are much more willing or likely to interweave professional talk with friendship. Women and girls use language as a means of maintaining a relationship, of feeling close and involved. Language is the glue that binds, and women are very concerned about and interested in what other women are talking about, even if (from a man's point of view) they are talking about nothing at all. In fact, one of the highest ratings of any television network production has been consistently obtained by an ABC daytime television show, "The View" in which a group of white and black women sit around and talk (Neilsen Ratings, May, 2010).
Men are more likely to focus on accomplishing something together, or talking about the accomplishments of others such as those who won a certain football game and those who excel or need improvement in their performance. Indeed, men are more likely to utilize speech as a means of maintaining or establishing status and to leave out details concerning their emotions or personal problems. Again, this can be directly attributed to the hunting way of life where skill and male vs male competitive (and cooperative) pressures not only determined success and failure but where weaknesses were ferreted out and ridiculed.
Women tend to become more interested in the details of a person's life and in discussing the details or their own lives as well as their thoughts and feelings, whatever they may be. When women are together this tendency naturally forms a bridge that maintains their rapport. They expect to be informed. However, when women are with men, they may feel that males is acting disinterest or not being communicative and is somehow lacking in the ability to express his feelings. Indeed, it is a common complaint expressed by my female patients and in the media; supposedly men do not express their feelings and are not as communicative, at least not in the company of the opposite sex.
In fact, often the man is disinterested and is wondering why she is talking about this or that and asking him all these "irrelevant" questions. Indeed, many men tend not to share their feelings and other personal details, because to do so might put them at risk with other men, who, being so competitive, may use it against them or make fun of them. Moreover, knowing that other men are not very interested in the social and personal aspects of another man's life, even when discussing family and children, men usually keep these topics quite brief as it seems irrelevant to the task at hand; to conquer the world, make more money, get that promotion, or to discuss those who are at present conquering the world, e.g. politics and sports (Gould, 1976). These are viewed as worthy topics to discuss at length.
Even when it comes to personal difficulties, men are more likely to keep it to themselves and answer in as few words as possible. By contrast, women are more likely to not only discuss these issues at length, but to discuss it again and again with all their friends.
Jake: "You know how it is."
Andy: "Yeah."
For many women, to keep these details to themselves is a breach of friendship and intimacy, for often their friends, and mothers insist on knowing the details and derive considerable enjoyment in hearing and discussing them. Again, consider Ms. Clinton revealing her husbands "emotional" problems, and "addictions" and "dysfunctions" and detailing his "emotionally abusive" childhood in a woman's magazine: TALK. Hence, women see nothing wrong or unusual about discussing or revealing intimate details about themselves or others, whereas men tend to fear they may lose power by doing so (as other men will make fun of them) and are thus so disinclined.
However, when it comes to plans, actions, and future goals, or even past conquests, the discussion between men is likely to be quite extensive, particularly if it can enhance their status.
Jake: "I'll say. You remember that blond that used to work at the...."
Girls and women, thus tend to focus on the social, supportive, familial and the communal aspects of interaction. Men tend to focus on the individual and the struggle for dominance over the community, the family, and each other. In this regard, when women interact they tend to be less confrontational as they are more sensitive to social and emotional nuances, and as one of their main interests is to maintain the cohesion of the group or the friendship. Indeed, these same exact sex differences are also evident among other primates including chimpanzees (De Waal, 1989), and are also directly related to sex differences in the division of labor over the course of evolution, and the differential male vs female pattern of neurological development.
Again, however, it is important to emphasize that there is much variability in these seemingly differential attitudes toward the use of language as a means of social and intimate interchange. Some men are more interpersonally inclined, and many gossip among themselves about sports, politics, and each other, although they do not call it as such. Similarly, many women are much more interested in the non-personal, political, business, informational aspects of communication and would rightly take extreme exception to the notion they spend their time talking predominantly about family, relationships, clothes, shoes, sex, or each other.
OVERVIEW: SEX DIFFERENCES, THE PARIETAL LOBE, CORPUS CALLOSUM & LANGUAGE
Superior female linguistic capacities appear to be related to their evolutionary history as child bearers, gatherers, tool makers, hide preparers and so on; tasks which would require superior social-emotional capabilities (e.g. child care, socializing) and which require considerable temporal-sequential motor capability and somesthetic sensitivity which in turn is associated with Broca's area and the parietal lobule in particular.
Gathering (and to a lesser extent tool making vs hunting), is far more likely to require bi-manual activation of the hands, and thus simultaneous activation of both parietal lobes and the motor areas of the frontal lobe. These activities might also be expected to coincide with increased functional and metabolic activity in the parietal lobes, which in turn might lead to an expansion of this tissue (inferior parietal) and an increased need to intercommunicate.
In this regard it is noteworthy that the posterior portions of the corpus callosum, i.e. the rope of fibers that interconnect the right and left parietal lobes, may be thicker and larger in females than males (Holloway et al. 1994). However contrary evidence abounds (reviewed in Holloway et al. 1994). Nevertheless, as gathering and related activities are more likely to involve both hands (vs hunting and throwing with a single hand), it might be expected that the posterior (female) corpus callosum would be larger because the female right and left parietal lobe were probably simultaneously activated and utilized by females more than males for the last 100,000 or so years. On the other hand, one might equally expect that the corpus callosal interconnections linking the motor areas, particularly those involved in fine motor functioning, would be more developed in females.
Given that women have clear language superiorities and unlike males were able to frequently and continually talk and chatter, if not with other women, then their children, sisters, or mothers, and as the IPL is involved in word finding, sentence construction, and organizing Broca's area for grammatical speech production, it might also be expected that the female IPL is more involved in language production as compared to males. Again, however, the same might be said of Broca's area.

It might also be expected that the female left parietal lobe would be more resistant to aphasic disturbances as compared to males whose language representation is more sparse and fragile. In fact, males are far more likely to become severely aphasic with left parietal injuries than females, who in turn more quickly recover from similar damage (Kimura, 1993). Conversely, females are far more likely to experience expressive aphasia with damage to Broca's area than males (Hier et al. 1994; Kimura, 1993). Again, however, they are also more likely to regain expressive speech functions.
On the other hand, it may be that sex differences in severity of initial language loss in anterior vs posterior lesions may be a function of sex differences in the vasculature and the origin and/or type of debri responsible for cerebral infarcts. In this regard, although females demonstrate clear language superiorities and have more neurological space in the right and left hemisphere devoted to linguistic and expressive functions (Joseph 1993), and in fact demonstrate bilateral activation when engaged in certain verbal tasks, whereas males tend to be more unilateral (Shaywitz, et al., 1995), the exact nature and neurological foundations for these differences are certainly still open to debate.
THE INFERIOR PARIETAL LOBE, APRAXIA, AND TEMPORAL SEQUENCING
The angular gyrus of the inferior parietal lobule (IPL) is unique to humans. An examination of the ape brain fails to find any evidence of this structure. The angular gyrus and IPL are crucially evolved in controlling temporal sequential hand movements including the manipulation of external objects and internal impressions (De Renzi and Lucchetti, 1988; Heilman et al., 1982; Joseph, 1982; Kimura, 1993; Strub and Geschwind, 1983). The evolution of the angular gyrus enabled humans to engage in complex activities involving a series of related steps, to create and utilize tools, to produce and comprehend complex gestures, such as American Sign Language, and to express and perceive grammatical relationships--capacities which are disrupted with lesions localized to the IPL (Joseph 1982, 1988, 1996, 2000a,b). In fact, the motor engrams that make possible temporal and sequential motor acts, including those involved in grammatical verbal expression, are partly localized within the IPL (De Renzi and Lucchetti, 1988; Heilman et al., 1982; Kimura, 1993; Strub and Geschwind, 1983). In fact, the IPL not only interacts with but appears to program the frontal motor areas for the purposes of producing fine motor, temporal-sequential movements, including the vocalization of speech units (Joseph 1982).
Those devoid of an angular gyrus/IPL, or those who have suffered a severe injury to this area, are generally unable to correctly manipulate or fashion complex tools -much less utilize them in a complex temporal sequence. This condition is referred to as apraxia; i.e. an inability to perform tasks involving interrelated steps and sequences (Joseph 1996). With severe IPL injuries, the individual may be unable to make a cup of coffee or put on their clothes, much less fashion or sew them together. Moreover, grammatical speech is disrupted and patients may suffer extreme word finding difficulty, or a conduction aphasia. That is, speech is no longer produced as Broca's area is disconnected from the IPL and Wernicke's area (Joseph 1982, 1996). Likewise, reading ability is disrupted as the IPL not only comprehends and produces gestures but visual symbols including written language. Hence, the IPL/angular gyrus (including the frontal motor areas) makes possible the ability to fashion and manipulate tools and organizes speech into vocabulary-rich, temporal sequential grammatical units.
As apes do not possess an angular gyrus, it appears that over the course of evolution, with the development of right handedness and selective pressures acting on gene selection across gathering/tool-making generations, the IPL/angular gyrus emerged as an extension of the auditory area in the temporal lobe and the superior parietal visual-hand area. Indeed, the parietal lobes are considered a "lobe of the hand" (Joseph 1982, 1993, 1996) and contain neurons which guide hand movements (Hyvarinen, 1982; Kaas, 1993; Lynch, 2010; Mountcastle et al., 1975, 2010) and which respond to visual input from the periphery and lower visual fields -the regions in which tool-making hands are most likely to come into view.
Because most individuals would use the right hand for tool making and the left for holding the tool, it is the left parietal lobe (which monitors the right lower visual field and controls the right hand) that guides and visually observes, learns and memorizes hand-movements when gathering, gesturing, or manipulating some object or constructing a tool. Over the course of evolution and as experience and the environment act on gene selection and induce neural plasticity, the parietal (and frontal) lobe expanded, the angular gyrus emerged, and neuroplastic alterations were induced in the adjoining motor-hand area in the frontal lobe including what would become Broca's speech area.
THE RIGHT HAND AND THE LEFT FRONTAL LOBE
Specifically, it is the lateral surface of the right and left frontal lobe which control vocalization and verbal-though production, the left frontal (Broca's area) producing the words, the right frontal the melody of language (Joseph 1982, 1986a, 1988a, 1996, 1999). The frontal lobes are interlocked with the association areas in the posterior regions of the cerebrum (Petrides & Pandya. 1999, 2001), including Wernicke's language comprehension area and the angular gyrus - inferior parietal lobe (IPL), as well as the memory centers in the temporal lobe (Joseph 1986a, 1996, 1999). The frontal lobes are therefore continually informed about and have continual access to information processed in these areas of the brain.


Specifically, the tissues of the mind known as Broca's Expressive Speech area, which is located in the left lateral surface of the frontal lobe (areas 45, 46), is responsible for programming the fine motor movements involved in the articulation of speech. Broca's speech area imposes temporal sequences on on the intonational contours of everything which is to be articulated so that the melodic features of generalized vocal expression come to be punctuated, sequenced, and segmented, and vowel and consonantal elements are produced (Joseph, 1982, 1992, 1996). Left hemisphere speech comes to be superimposed over limbic and right hemisphere melodic language output (Joseph 2000). Thus, the primary area representing the oral-laryngeal musculature, is programmed by Broca's expressive speech area which becomes active prior to vocalization and during subvocalization as indicated by functional imaging.
LEFT FRONTAL DEVELOPMENTAL DOMINANCE FOR MOTOR FUNCTIONS
The motor areas of the frontal lobe include Broca's expressive speech area, Exner's hand area, the supplementary motor area, and the secondary and primary motor areas. There is a one-to-one correspondence between single neurons in the primary motor area and single muscles such that the musculature of the entire body surface is represented according to their importance (Chouinard & Paus 2006; Dum & Strick 2005; Verstynen, et al. 2011). For example, the hands, fingers, face and mouth, have a greater representation than the musculature of the back. The representation of the musculature is more diffuse in the secondary premotor area. The premotor transmits its motor impulses to the primary motor areas and subcortical regions of the brain but receives its marching orders from the supplementary motor area (Joseph 1999b; Nachev et al. 2008).
Movement, including those involved in expressive speech, is made possible by ropes of descending axons nerve fibers which project from the frontal motor areas to the brainstem and spinal cord. These nerve fiber ropes are referred to as the pyramidal and corticospinal tracts (Joseph 1999n; Miller & Cummings 2006; Risberg & Grafman 2006). Hence, if the motor areas of the right or left frontal lobe are destroyed, the left or right half of the body becomes paralyzed. However, if damage is restricted to Broca's area, the ability to speak in words is lost.
Among 80% of humans, the motor cortex of the left frontal lobe is dominant for expressive speech, and for controlling handedness and programming gross and fine motor, temporal-sequential, movements of the musculature, and of the right hand and fingers in particular . It is this dominance and the functional organization of the left frontal lobe and the left hemisphere, which led to the evolution of modern human language (Joseph 1982, 1990, 1996, 2000a), the expression and comprehension of which is also associated with the functional integrity of the left half of the brain.
Left hemisphere dominance is due to factors associated with fetal development and experiential influences on gene expression over the course of evolution.
During fetal development, and due to the earlier maturation of the left cerebral motor areas, the descending corticospinal tract from the left hemisphere matures and establishes its connections with the brainstem and spinal cord in advance of the axons from the right hemisphere (Joseph 1982). The left corticospinal tract grows more quickly and descends into the brainstem, and then crosses at the pyramidal decussation, and then descends into the spinal cord in advance of those fibers from the right (Kertesz & Geschwind 1971; Yakovlev & Rakic 1966). This provides the left hemisphere and the right hand with a competitive advantage in establishing both motor control and thus right hand dominance. Thus, the left hemisphere motor areas gain a competitive advantage over the right hemisphere, and acquire greater motor control over brainstem and spinal motor neurons controlling movement (Joseph, 1982). Therefore, the somatomotor areas of the left frontal lobes mature at a more rapid rate than the right frontal lobes and by the first year tissues such as Broca's area overtake their right sided counterparts (Scheibel, 1991, 1993). However, because the corticospinal tract does not become functionally mature or myelinated until the 8th-12th postnatal month (Yakovlev & Lecours, 1968) it is only as the infant approaches their first birthday that handedness become obvious or apparent. Corticospinal and pyramidal axons continue to become increasingly well myelinated over the course of the second year (Debakan, 1970; Langworthy, 1937; Yakovlev & Lecours, 2007)--a process which continues into late childhood (Paus et al., 1999). Hence, around the 8-12th month the neocortex and frontal motor areas begin to increasingly exert hierarchical control over the limbic forebrain, brainstem, and musculature, and most individuals are right handed.
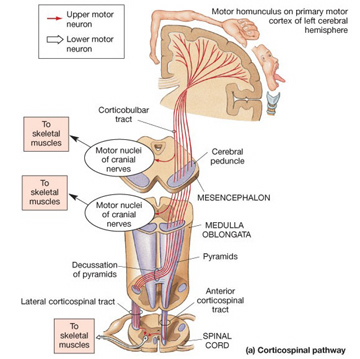
Broca's expressive speech area appears to be an evolutionary partner, if not an outgrowth of the adjacent frontal area controlling fine motor movements of the hands and fingers; referred to as Exner's writing area and which is located near the foot of the second frontal convolution of the left hemisphere, occupying the border regions of Broadmans areas 46, 8, 6. Exner's area appears to be the final common pathway where linguistic impulses receive their final motoric stamp for the purposes of writing; i.e. the formation of graphemes and their temporal sequential expression. Thus, Exner's writing area appears to program the ajacent hand-area represented in the primary motor areas (e.g. Boroojerdi et al., 1999) so that lingusitic impulses received through Broca's area, can be integrated into hand movements so that words can be written down.
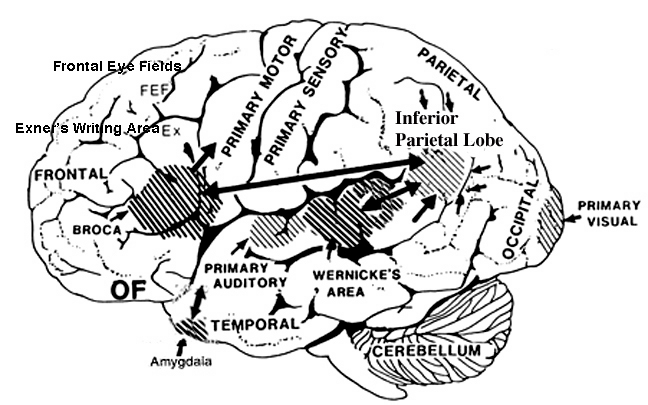
Exner's area is dependent on Broca's expressive speech area with which it maintains extensive interconnections. In fact, Exner's writing center extends to and appears to become coextensive with Broca's area (Lesser et al. 1984)--which in turn was originally a hand area--at least in primates. Broca's area possibly acts to organize and relay impulses received from the posterior language zones to Exner's area in instances where written expression is desired. Exner's area, in turn, transfers this information to the secondary and primary motor areas for final expression.
Because these areas are linked, when most people talk, their expressive speech is often accompanied by hand movements. Conversely, whereas damage to Broca's area will significantly impair the ability to speak in words, destruction of Exner's area results in apraxia.
Broca's and Exner's areas do not exist in the brains of apes. In fact, the frontal lobes are not well developed in these species, or in ancestral pre-human species, be it Australopithecus, H. habilis, H. erectus, or Neanderthal (Joseph 1993, 1996, 2000b). Rather, the frontal lobe does not significantly expand in size until the onset of the Upper Paleolithic and the evolution of the Cro-Magnon people, complex tool making technology, and language (Joseph 1993, 1996, 2000b).
The primary auditory receiving areas are located in the superior temporal lobe. Once received and processed, these auditory signals are transferred to the immediately adjacent Wernicke's area which associates these sounds and comprehends the words of language; whereas the the auditory association area in the right temporal lobe comprehends environmental sounds and melodic and emotional vocalizations (Joseph 1982, 1988a, 2000). Via a massive neural pathways (the corpus callosum) the right and left auditory receiving areas work together when presented with complexity and paralinguistic features which require analysis (Just et al., 1996; Michael et al. 2001; Schlosser et al., 1998).




Movement and motor functioning are dependent on the functional integrity of the basal ganglia, brainstem, cerebellum, cranial nerve nuclei, the motor thalamus, spinal cord, as well as the primary, secondary and supplementary motor areas of the frontal lobes. These areas are all interlinked and function as an integrated system in the production of movement.
For example, Exner's writing area is in part, within areas 6 and becomes active prior to (as well as during) hand movements and appear to program hand movements, whereas the frontal eye fields (within areas 6,8,9) becomes active prior to (as well as during) eye movements and appears to program eye movements.
The motor areas do not just program, but anticipate, as well as memorize certain movement programs. In fact, individual neurons within the motor cortex (of monkeys) have been shown to learn and neurally encode serial order when performing a context-recall visual-memory scanning task (Carpenter et al., 1999). Moreover, neural activity will change when the serial order is changed and in accordance with the direction of movement as well as stimulus position and movement (Carpenter et al., 1999; Passingham, 1997). In fact, according to Carpenter et al., (1999, p. 1755), although "individual cells prove largely independent information about the items in the sequence... during different epochs of presentation of the stimuli, different patterns of distributed activity in even small ensembles of motor cortical cells are sufficiently distinct and robust to provide a basis for encoding the sequence," which in turn allows for memory scanning and retrieval.
In addition, "motor" neurons in the inferior-ventral portion of the (monkey) premotor cortex can code for the spatial location of nearby sounds, particularly those originating near the head (Graziano et al., 1999). Indeed, these neurons appear to be "multi-modal" and can also respond to visual and tactile stimuli that impinge near or upon the face (Graziano et al., 1999).
However, these motor areas also act in association with visual-spatial information received from the parietal and occipital lobe, so that the movements of the body, the hands in particular, can be guided while moving about in visual space. In this way, gross movements of the hand and arm can also be coordinated with visual impressions, thereby enabling an individual to throw an object with accuracy, such as a spear to impale a running animal.
Visual information is transmitted to the frontal eye fields (FEF) which are located along the superior lateral convexity of the frontal lobe and which are part of the premotor area and encompasses all of area 8 as well as portions of areas 9. As the name implies, the FEF is concerned with eye movements, and appears to program the corresponding primary areas so as to guide gaze shifts and visual tracking. The FEF receives visual information from the primary and association visual cortices in the occipital lobe (17, 18, 19), the auditory association (22) and multimodal visual association areas (20) in the temporal lobe (Barbas & Mesulam, 2011; Jones et al., 1978; Jones & Powell, 1970), and the somatosensory association area (Crowne, 1983). It also shares interconnections with the caudate, superior colliculus, and oculomotor nucleus (Astruc, 1971; Knuzle & Akert, 1977; Segraves & Goldberg, 1987). Hence, the FEF receives information concerning the auditory, tactual, and visual environment and is multimodally responsive.
The FEF coordinates and maintaines eye and head movements, gaze shifts, and thus orienting and attentional reactions in response to predominantly visual, but also tactile and auditory stimuli. It is also involved in focusing attention on certain regions within the visual field, particularly the fovea guiding eye movements while reading, writing, and when tracking and throwing.
In addition to visual tracking and supporting focused visual attention, neurons in the FEF demonstrate anticipatory activity; that is, firing before a response is made. In fact, these neurons will continue to fire at a high rate until the behavior is initiated. In this regard, neurons in the FEF probably exert a countering influence so that attention does not drift and attention and eye position remains stabilized and focused on the primary retinal target.
Moreover, cells in the FEF will fire slectively in response to stationary and moving stimuli, to objects which are within arms reach, as well as to tactual stimuli applied to the hands and/or mouth. In fact, as an object approaches the face and mouth, some of these cells correspondingly increase their rate of activitity (Rizzolatti et al., 2011ab). Hence, cells within the FEF are highly involved in mediating sustained attention and orienting reactions of the head and eyes, maintaining visual fixation and modulating visual scanning, as well as coordinating eye-hand and hand to mouth as well as smooth pursuit eye movements.
Motor functioning and movement, however, is also dependent on the parietal lobes, thalamus, basal ganglia, brainstem, cerebellum, and spinal cord, structures which are directly or indirectly interconnected (Kaas, 1993; Mink, 1997; Passingham, 1997; Schmahmann, 1997).
For example, smooth, purposeful, coordinated movement requires sensory feedback which is provided by the primary and association/secondary somatosensory areas located in the parietal lobe and which contain neurons which guide the hand and arm in visual space. These cortices are intimately linked with the primary motor and the motor association/secondary and supplementary motor areas as well as the basal ganglia, brainstem, and cerebellum--regions which become highly active during and often prior to movement initiation (Passingham, 1997). For example, when human subjects learn a sequence of finger movements, functional imaging reveals increased activity in the primary and premotor cortex, and the primary and association somesthetic cortex, as well as in the cerebellum, striatum, ventral thalamus and cerebellum (Passingham, 1997). In fact, most of the "motor" area also contain independent motor maps of the body, which is why, if a subject moves an arm, leg, or shoulder, each of these areas becomes active almost simultaneously--at least as demonstrated with functional imaging.
BROCA'S EXPRESSIVE APHASIA
Broca's speech are is located in the general vicinity of the posterior- inferior region of the left frontal area (i.e. third frontal convolution), and includes portions of areas 45, 6, 4, and all of area 44. This region is multimodally responsive (Passingham, 1997) and receives projections from the auditory, visual, somesthetic areas (Geschwind, 1965; Jones & Powell, 1970), as well as massive input from the inferior parietal lobule and Wernicke's area via a rope of nerve fibers referred to as the arcuate fasciculus (which also links these areas to the amygdala). In addition, Broca's area receives fibers from and projects to the anterior cingulate as well as the brainstem periaqueductal gray which subserves vocalization.
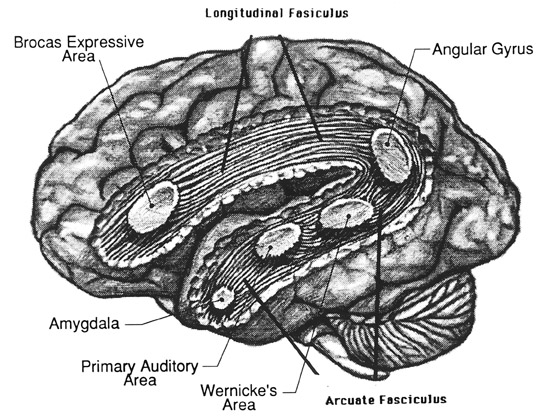
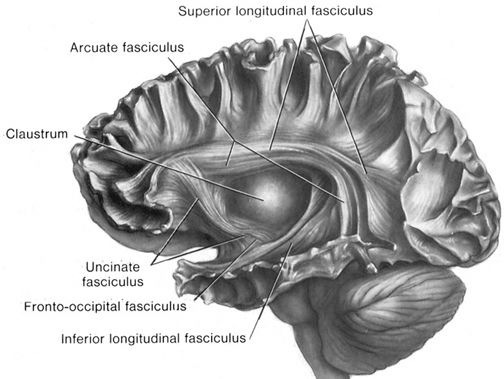
The importance of Broca's area and the left frontal lobe has also been demonstrated through functional imaging. For example, the left frontal lobe becomes activated during inner speech and subvocal articulation (Paulesu, et al., 1993; Demonet, et al., 1994). The left frontal lobe also becomes highly active when reading concrete and abstract words (Buchel et al., 1998; Peterson et al., 1988), and when engaged in semantic decision making tasks (Demb et al., 1995; Gabrielli et al., 1996). Moreover, activation increases as word length increases and in response to long and umfamiliar words (Price, 1997).
With injuries to Broca's area the individual loses the capacity to produce fluent speech. Output becomes extremely labored, sparse, and difficult, and they may be unable to say even single words, such as "yes" or "no". Often, immediately following a large stroke patients are almost completely mute and suffer a paralysis of the upper right extremity as well as right facial weakness (since these areas are neuronally represented in the immediately adjacent area 4). Patients are also unable to write, read out loud or repeat simple words. Interestingly, it has been repeatedly noted that almost immediately following stroke some patients will announce "I can't talk", and then lapse into frustrated partial mutism.
With less severe forms of Broca's (also referred to as expressive, motor, nonfluent, verbal) aphasia, speech remains labored, agrammatical, fragmented, extremely limited to stereotyped phrases ("yes", "no", "shit", "fine") and contaminated with syntactic and paraphasic errors i.e. "orroble" for auto, "rutton" for button. Writing remains severely effected, as are oral reading and repetition. Such patients also have mild difficulties with verbal perception and comprehension (Hebben, 1986; Maher et al. 1994; Tramo et al., 1988; Sarno, 1998; Tyler et al. 1995), including the ability to follow 3-step commands . Commands to purse or smack the lips, lick, suck, or blow are often, but not always, poorly executed; a condition referred to as bucal-facial apraxia. Non-speech oral movements are seldom significantly effected.
With mild damage, patients may demonstrate severe confrontive naming and word finding difficulties (anomia), as well as possible right facial, hand, and arm weakness. Speech is often characterized by long pauses, misnaming, paraphasic disturbances and articulatory abnormalities . Stammering and the omission of words may also be apparent. Similarly, electrical stimulation of this region results in speech arrest (Ojemann & Whitaker, 1978; Lesser et al., 1984) and can alter the ability to write and/or perform various oral-facial movements. It is noteworthy that even with anterior lesions or surgical frontal lobectomy sparing Broca's area a considerable impoverishment of spontaneous speech can result (Luria, 2010; Milner, 1971; Novoa & Ardila, 1987). Disturbances involving grammar and syntax, and reductions in vocabulary and word fluency in both speech and writing have been observed with frontal lesions sparing Broca's area. In word fluency tests, however, simple verbal generation (e.g. all words starting with L) is usually more severely impaired than semantic naming, e.g. all animals which live in the jungle--which is presumably a function of semantic processing being more dependent on posterior language areas.
THE EVOLUTION OF THE SPEECH AREAS
Non-human primates lack a functional Broca's area. Among non-human primates, the left frontal lobe, including the tissues homologous to Broca's area, does not subserve speech or vocalization (Jurgens et al. 1982; Myers 1976). Vocalization in apes and monkeys has a non-segmented organization, consisting of moans, screams, barks, grunts, pants, and pant-hoots (Erwin, 1975; Fedigan, 1992; Goodall, 1986, 1990; Hauser, 1997) and is the province of the limbic system and brainstem; e.g. the cingulate gyrus, amygdala, hypothalamus, and periaqueductal gray (Joseph, 1982, 1993; Jurgens, 1990; MacLean, 1990; Robinson, 2007; 1972). Therefore, although damage to Broca's area in humans results in a profound expressive aphasia, similar destruction in non-human primates has no effect on vocalization rate, the acoustical structure of primate calls, or social-emotional communication (Jurgens et al. 1982; Myers 1976). Rather, in primates, those tissues homologous to "Broca's area" are directly involved in motor activity of the arms and hands (Rizzolatti, et al. 1988). Moreover, neural pathways linking the primate IPL with the homologous primate "Broca's area" are only weakly developed (Abolitiz and Garcia, 1997). Apes and monkeys, however, do not possess an angular gyrus.
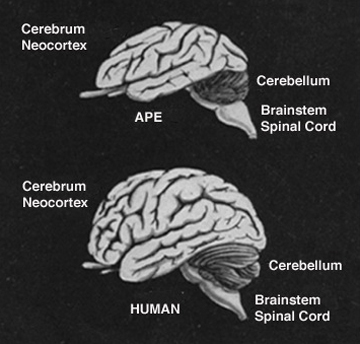
The angular gyrus sits at the junction of the posterior-superior temporal and the occipital-parietal lobes, and is critically involved in naming, word finding, grammatical speech organization, and is in part an extension of and links Wernicke's with Broca's area (Joseph, 1982, 1999e; Goodglass & Kaplan, 2000). Through its extensive interconnections with the adjacent sensory association areas, the IPL/angular gyrus receives and assimilates complex associations, thereby forming multi-modal concepts, and acts to classify and name this material which is then injected into the stream of language and thought. The IPL/angular gyrus, in concert with Wernicke's area, transmits this information to Broca's speech area, which in turn organizes the immediately adacent oral, laryngeal motor areas (Foerster, 1936; Fox, 1995; Joseph, 1992, 1999a).
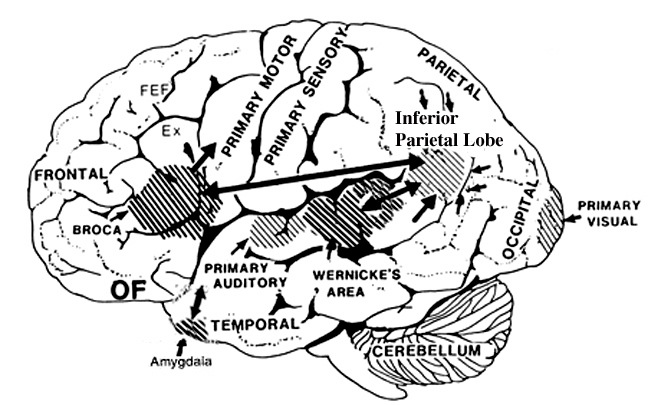
Chimpanzee Brain
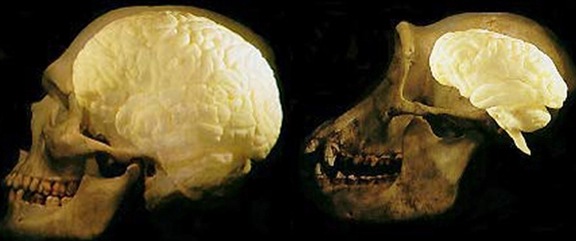
Human (Left) and Chimpanzee (Right) Brain
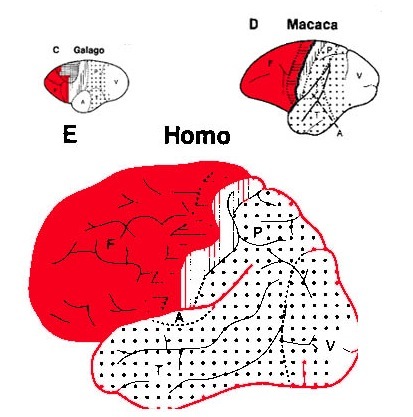
Presumably, left planum temporal and hemisphere dominance for vocal perception and comprehension gradually increased in the transition from Australopithecus, to H. habilis, to H. erectus, to Neanderthals. As first proposed and detailed elsewhere (Joseph, 1993, 1996, 2000b) as Wernicke's area, the parietal-hand areas and the IPL expanded, merged, and collectively gave rise to the angular gyrus, auditory input began to be sequenced, and Wernicke's area became specialized for perceive and comprehending language units.
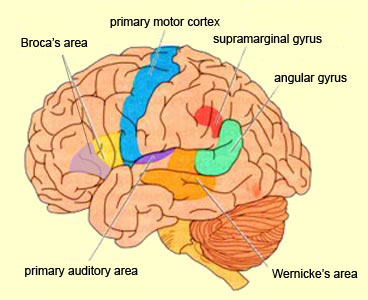

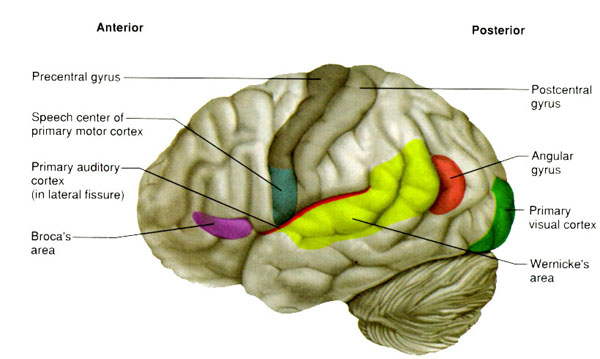
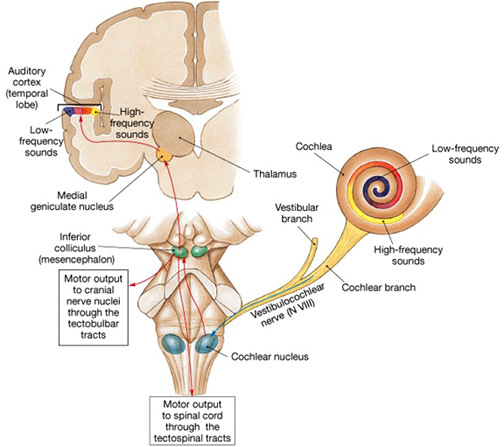
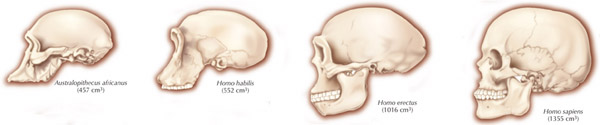
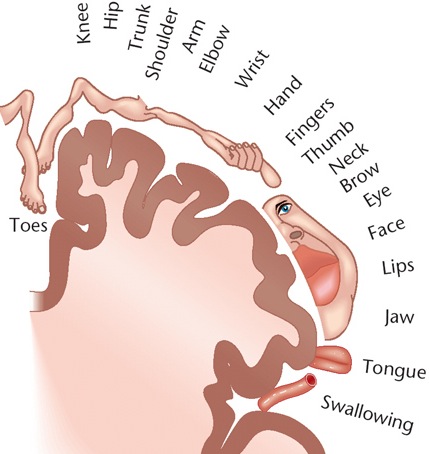
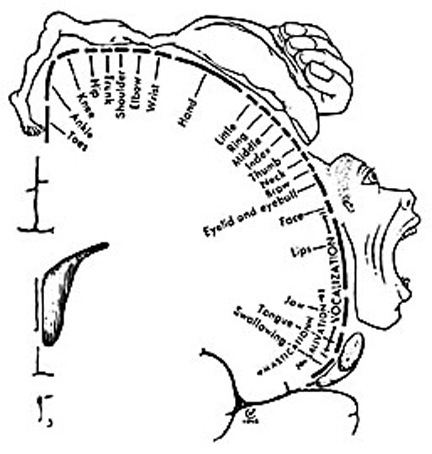
Hence, Wernicke's area began sequencing auditory input, and Broca's area was transformed from a hand area to a speech area and ceased to control hand movements whereas Exner's area became specialized to control the fingers and hands. Once Broca's area evolved into a speech area, it began to organize the adjacent primary motor oral-laryngeal areas so as to express the words and sentences transmitted from the IPL and Wernicke's area.
Hence, in addition to manipulating tools in a temporal sequential fashion, the evolution of Exner's area and the IPL/angular gyrus enabled humans to manipulate linguistic impulses in temporal sequences which were transmitted to the oral-laryngeal musculature which began to vocalize units of speech.
However, as based on an analysis of tool technology, it can be concluded that Australopithecus, H. habilis, H. erectus, and Neanderthals did not possess the neurological sophistication for vocalizing complex human language, and had not yet evolved an angular gyrus or a functional Broca's area (Joseph 1992, 1993, 1996, 2000a,b). Rather, the evolution of modern speech likely corresponded to the evolution of the Upper Paleolithic female gatherer and tool maker.
THE FEMALE BRAIN & THE UPPER PALEOLITHIC
The frontal motor area representing the hand is immediately adjacent to and intimately interconnected with the primary motor areas mediating oral, laryngeal, and mandibular movements, including Broca's area. Hence, manual activity, right handedness and expressive speech are directly related (Bradshaw and Rogers 1992; Corbalis, 1991; Joseph, 1982; Kimura, 1993), which is why when speaking, humans commonly gesture with the hands, the right hand in particular. However, it was not until the late Middle Paleolithic that up to 90% of Paleolithic Humans may have become right handed (Cornford 1986). Similarly, it was not until the Middle to Upper Paleolithic transition, 35,000 years ago, that tool making became literally an art and complex multifaceted features were incorporated in their construction and utilization (Chauvet et al., 1996; Leroi-Gourhan, 1964, 1982).
The Middle/Upper Paleolithic transition is characterized by the creation of complex bone tools, the sewing needle, and personal adornments such as carefully shaped beads of bone, ivory and animal teeth, animal engravings, perforated shells, statuettes, drawings, and paintings of animal and female figures (Chauvet et al., 1996; Clark, 2007; Leroi-Gourhan, 1964; 1982).
As the creation and wearing of personal adornments, and domestic tool construction and use is associated with the human female, and as tool making and gathering often involves both hands (albeit the right more than the left), it might be expected that the human female frontal-parietal areas may have functionally evolved in a manner different from men. That is, both the left and right half of the female brain may have become organized for producing motor sequences and grammatically complex vocabulary rich speech.
Moreover, as the parietal lobe and IPL/angular gyrus act on and program the frontal motor and speech areas, whereas the left frontal areas adjacent to the hand and face area control and program speech and hand-related fine motor activities (Joseph 1982, 1996, 2011d), it might be expected that sex differences would be more pronounced in Broca's and the parietal areas. In fact, Broca's area appears to be larger in the female.
Moreover, the posterior corpus callosum which interconnects the right and left parietal lobes, appears to be significantly larger in women than men (see Holloway et al., 1993, for evidence pro and con), whereas language appears to be represented in the right half of the female to a greater degree than it is represented in the right half of the male brain (Bradshaw et al., 1977; McGlone, 2010; Shaywitz et al., 1996) --a sex difference which may also account for her superior language capabilities but also her relatively inferior visual-spatial abilities (due to functional crowding). Left hemisphere dominance for verbal functioning and right hemisphere language (and greater emotional language) representation coupled with the enhanced ability of the right and left female-IPLs to communicate via the corpus callosum, may account for why women (versus men) are less likely than men to become aphasic with a left parietal lesion (Kimura, 1993; Mateer et al., 1982); i.e. women have language-related brain tissues in reserve and are able to continue talking so long as Broca's expressive speech area is uninjured.
SUMMARY & CONCLUSIONS
For much of human evolution females have engaged in tasks promoting, requiring, and involving rapid temporal sequential bilateral fine-motor skills such as gathering and domestic-tool construction and manipulation. Given an innate tendency to vocalize more than males, these activities, coupled with prolonged child care and mutual child-mother vocalizations, gave impetus to the evolution of the neocortical speech areas and a female language (and social emotional speech) superiority.
In contrast to gathering groups in which females could vocalize with each other and their young, males instead pursued their own violent tendencies and became silent hunters of big game. Over the course of evolution the male proclivity to silently travel long distance in the pursuit of prey exaggerated an already innate male visual-spatial perceptual superiority.
Nevertheless, although hunting does not promote the evolution of speech, males also acquired language skills through maternal genetic inheritance and and as he had a mother who would talk to him and teach him language. Thus, like the proverbial Eve, woman the gatherer provided man the hunter with the fruit of linguistic knowledge and what would become grammatically complex, vocabulary-rich speech, language, and linguistic consciousness.
FOOD, SEX & THE BIG BRAIN
Foraging, scavenging, and the chasing and hunting of small game has probably been a dominant activity of human beings and their ancestors for several million years. Naturally, our mind and brain has been tremendously influenced by these activities and has evolved accordingly.
Initially, both the male and the female of the species probably engaged almost equally in scavenging, gathering and the hunting of small game until just a few hundred thousand years ago with the onset of big game hunting. It was possibly this event, coupled with the rapid and progressive development of mans big brain several thousand years earlier, that a divergence in the mind of man and woman appears to have increasingly occurred. The hunting of large game animals appears to have become the dominant domain of the male, although they probably continued to assist in gather on occasion. Presumably females continued to predominantly engage in gathering, and to a lesser extent fishing and the hunting of small game, and only occasionally assisted males during the hunting and stalking of large animals.
Nevertheless, with this general division of labor, one might suppose that over the course of several hundred thousand years, tremendous differential influences on mental and brain functioning resulted due to selective evolutionary pressures on survival. That is, those who were best able to adapt were most likely to pass on their genetic traits to the next generation. In consequence, the brains and minds of men and women probably became adapted and molded accordingly each in accordance with the activities they were best at.
In this regard, as gathering and the harvesting of wild foods involves perseverative, temporal-sequential hand movements, those who were most successful at these activities probably had brains that were best adapted to these tasks. However, if the right hand is being used predominantly for picking and gathering, whereas the left hand is more likely to hold a recepticle, one might suspect that the left half of the brain would become even more adapt at temporal-sequential processing.
In contrast, as the searching and stalking and killing of prey would require good visual-spatial skills and sensitivity to environmental and non-verbal nuances, those who were most successful at these activities would in turn continue to develop, and pass on to the next generation, these same capabilities.
Over the course of human evolution, the human brain and the human head became increasingly larger. The progressive and significant increase in brain size, in turn required an adaption in the pelvic opening of these ancient females. A bigger brain comes in a bigger head. Hence, eventually the hips of the human female became wider as well so as to accommodate a larger brained baby. With a wider pelvic opening and with wider hips, a big brained baby could be delivered without becoming damaged or lodged inside the womb unable to emerge. If that were to happen, the baby, the mother, or both would die.
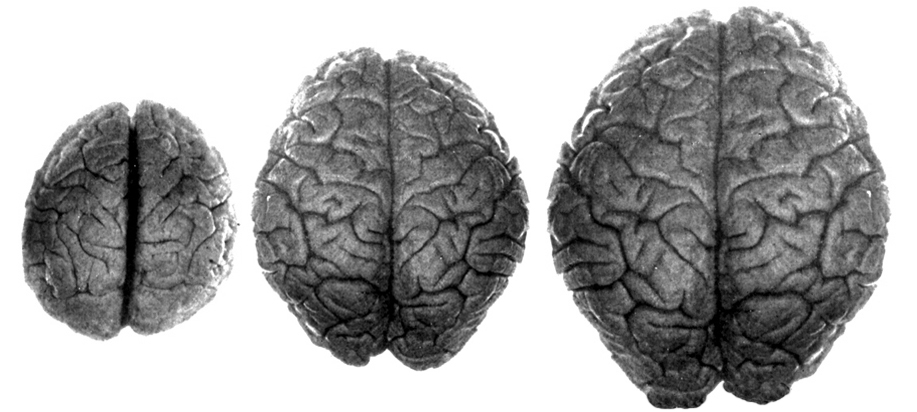
Comparative Brains: Australopithecus, H. erectus, Modern Humans
The human brain, of course is not fully developed at infancy and in fact is only about a third of the size of an adult. If it was too close to the adult size it would be impossible for an infant to be born. In consequence, the brain has to do a tremendous amount of growing after birth, a process that continues unabated for the first 10 years of life and then slows over the course of each ensuing decade.
However, by prolonging infant development the period of helplessness became lengthened as well which in turn necessitated prolonged child care. Hence, in contrast to most mammals and all other forms of life which remain helpless and unable to fend for themselves for only a few months at most, humans remain helpless for 4-6 years. This is in fact 4-6 times longer than for monkeys, and twice as long as for all other apes, which, like their female counterparts, invest a considerable amount of time in childcare.
Thus, the human infant, including an infant Cro-Magnon, Neanderthal, and even Homo erectus, required almost the full time presence of a mother for many years. This increased dependency in turn reduced female mobility as well. Hence, at a minimum, human females have been engaged in prolonged child care for at least half a million years or longer.
In consequence, having a baby or several children to watch over, and being unable to run as fast and being more clumsy in the attempt, the adult human female did not make for as good and as reliable hunter as compared to men. You can't carry an infant while stalking prey because your hands and arms must be free and you need quiet. The sudden cooing or crying of an infant is likely to result in an empty stomach. Hands and arms must also be free to carry weapons. Hence, there were a number of forces acting on females which conspired against their ability to excel as big game hunters. Indeed, her smaller stature also made her a less formidable foe and possibly even a a tempting prey.
Hence, for good reasons females probably tended to wander less than males, to stay closer to the home base, and to engage in activities compatible with a lifestyle that included the nurturance of infants for years at a time. The ultimate consequence of these particular limitations was that the human female probably remained more involved in gathering and the occasional hunting of small game closer to the home base whereas males were more free to engage in new activities such as big game hunting.
Again, one might naturally assume that to be successful as a hunter required certain adaptive changes in the human brain, alterations which would be maximal among males. Conversely, gathering, an activity involving prolonged and rapid temporal and sequential hand movements, would in turn exert its maximal selective pressures on the brain of the female. That is, those who were best at these activities were more likely to survive and pass on their genetic contributions and thus their adaptive skills, to their progeny.
Although a woman, or a band of women, could probably live and flourish without the aid of the bigger and more muscular male, and certainly could eat quite well without ever savoring a steak from a big game animal, her smaller size and the children that she bore, certainly put her at a disadvantage when dealing with males. Indeed, since our ancestors may have been slaughtering each other since the time of Australopithecus, one might speculate that it may have certainly been to her advantage to have at least one male around to aid in her protection as well as the rearing and education of the children. That is, she may have needed a male to protect her from other males who might rape or kill her and her children. Although perhaps highly debatable, there were probably other advantages to have a man about the house (or cave), as well such as being able to partake in the high protein diet that meat afforded.
In part, this bonding of a man to a woman, was aided by the establishment of a base camp and the maintenance of a family social group, events which appear to have first had their onset during the time of Homo Erectus (who lived from 1.9 million to about 300 thousand years ago). However, from an evolutionary perspective it was necessary to provide the man with some incentive to return home and thus maintain the sanctity of the family. The solution was possibly an appeal to the limbic system: food and sex.
To insure that the male return to base camp and share his spoils with her and her infant required that males become bonded to females or at least highly attracted so as to return again and again. The manner in which this, at least in part, may have been achieved was for females to become sexually receptive and sexually available 24 hours per day, 365 days a year. Indeed, the human female is the only female regardless of species, who is able to have sexual intercourse during times when she is not biologically receptive in order to become pregnant. For all other species, sex serves as a means of reproduction. Among humans, sex is a source of pleasure and serves as a means of bonding a man and a woman.
As is well known, for most other mammals there are long periods in which females are not sexually receptive and this is characterized by long period of non-sexual activity in which males show little or no interest in females. When she becomes sexually receptive there is a short period of frenzied sexual activity in which males will literally go sexually crazy for days until the female ends her estrus or "heat." of course, when a female goes into heat all other activities cease or greatly suffer.
It may have been sometimes during the rein of Homo erectus that this great change may have occurred and the female estrus disappeared altogether. Indeed, it was during the time of Homo erectus (or his immediate ancestor) that the first base camps also came into existence. However, even if we assume that this sexual revolution did not come until the end of their rein, that is, around the time period in which the first primitive Homo sapiens arrived on the scene it would still appear that full time sexual availability had it's onset somewhere between 300,000 and 500,000 years ago. Nevertheless, in consequence of that momentous occasion, the human female has been the sexiest of all other females ever since.
Her becoming sexually available at all times enabled man and woman to become free of the purely biological and hormonal influences which drive and control sexual behavior. They could now postpone sexual activity and could now decide for themselves when and if they were going to have sex, as well as where it would take place. Moreover, unlike the apes who only on occasion and for brief time period can enforce a selective preference, humans were also able to decide with whom they were going to have sex and could enforce their preference via denial and through bonding.
All in all, with the complete availability of a full time sexual partner coupled with the ability to make personal choices and to act on preferences, human sexual relations became more enduring and so to did the relationships between men and women; all of which probably promoted the development of language and the art of conversation.
Man, still functioning at the behest of his limbic system, could now have one almost insatiable need repeatedly satisfied. However, as noted, a second insatiable need, food, was also provided by the female. That is, gatherers provide the bulk of the food supply. There were thus two very good reasons for males to not only come home, but to stay home for prolonged time periods as females were not only providing for their sexual needs, but gathering and probably preparing their supper as well. Man was becoming domesticated.
FOOD & THE ORIGINAL "BREAD WINNERS"
With the exception of those who lived in the very coldest of climates where a gathering way of life would be quite difficult due to the scarcity of edible vegetable matter, for at least the last 100,000 years until perhaps about 10,000 years ago, females, and not males, appear to have been the main providers of food. Big and even small game hunting has always been (except in the much colder far northern climates) a supplementary means of acquiring an adequate food supply.
Indeed, even among the great majority of the very few modern hunting and gathering societies in existence today, spoils from hunting account for only about 35% of the diet. In contrast, gathering, which, we are assuming, has been the predominant domain of the female for the last 300 or so thousand years, accounts for the remainder. Indeed, even among the Cro-Magnon, where hunting was the center of religious and artistic life, 60-80% of their diet consisted of fruits, nuts, grains, honey, roots and vegetables, which was probably gathered by the females. Hence, it probably was the female, and not the male who probably wielded economic dominance for a significant period of our history.
Indeed, given woman's dominant role as gatherers and since they were probably generally responsible for both reproductive and most subsistence activities, males could be almost be considered a co-producing dependent. That is, women were probably the original "bread winners." Perhaps it is this ancestral tendency to nurture and provide that enables some modern women to not only tolerate but to support men who simply sponge off them.
Not surprisingly, males recognized that females were a tremendous economic asset, and in many societies, dominant males would try to accumulate as many wives or lovers as possible. However, although sexual availability may have played some role in this procurement during ancient times, the fact that females were the dominant producing partners meant that with many women, the man would be freed from having to spend all his time engaged in the pursuit of food, its preparation or even the maintenance of dwelling. The more females, the more leisure time and thus the more time to engage in recreational and artistic tasks.
Indeed, such a relationship, at least for dominant males, may have been rather ideal. For the first many years of their lives they were provided for by their mothers, and then later in life they were cared for by their wives. On the other hand, one could argue that not only were women the main providers, they were being exploited for the last 100,000 years as well.
Nevertheless, with increased leisure time and the evolving complexity of social relations, tremendous selective pressures were no doubt brought to bare on the mind and brain of human beings. Only those who could selectively adapt to these changing conditions could pass on their traits and genes. Not only were the right and left brain evolving new functional capabilities associated with sex differences in socializing, gathering and big game hunting, but correspondingly, about 50,000 years ago, the frontal lobes of the brain tremendously expanded in size. With the development of the frontal lobes came the capacity to engage in long term planning, the formation of goals, and the ability to maximally inhibit one's immediate desires and impulses. This great change came about with the appearance of the Cro-Magnon and social relations expanded beyond small bands to large tribes.
HUNTERS: BRINGING HOME THE BACON
Since food gathering was such an extremely important part of economic existence for so many hundreds of thousands of years until very recently, it might be asked, why has so much emphasis been placed on the importance of man the hunter and along with it, the false notion that he was the main provider?
To better understand the role of providing meat in the assumed dominance of man as the most important provider, let us consider our nearest living evolutionary cousins, the apes. When a chimpanzee captures another living creature for the purpose of consumption others will rapidly and excitedly gather around and beg and beg for just a morsel of the meat. Nothing like this happens when they forage for vegetables or insects. Indeed, the hunting and capturing of meat has an immediate highly arousing excitatory effect on the whole band all of whom will gather around in hopes of being given a tiny morsel.
Moreover, Chimps and baboons will chase after and kill small animals but they do not eat dead ones. That is, they do not scavenge. For them the pleasure of the meat is tied to the hunt, capture and killing. However, it is the male who predominantly engages in these acts.
Thus among the chimpanzees, the procurement and eating of meat for some reason promotes considerable social excitement and food sharing. In contrast, they do not share and show no interest in sharing or receiving any of the vegetable matter that another chimp may have found and is eating.
Similarly, among hunter gatherer societies, the procurement of meat promotes food sharing and the eating of meat is thus a very social activity. Indeed, when the hunter returns he is likely to be met by most members of his band all of whom will eagerly seek a share of the spoils. The hunter will naturally give the largest pieces to his own relatives and the smaller shares to friends and co-band members. These individuals in turn will give a share of what they receive to their own special friends and relatives. Hence, a certain degree of group cohesion and bonding occurs when meat is caught and shared and the sharing reinforces the social bond. Meat thus becomes an important currency of exchange.
In contrast, when females bring home the vegetable matter that they have gathered, there is no excitement and there are no begging hands eagerly seeking a share. What the female procures is shared only with her own immediate family.
Indeed, this same emphasis on the importance of meat can be found repeatedly in Genesis and the Books of Moses, as God demanded his sacrifices to be of living flesh. Hence, when Abel offered God "the firstlings of his flock, the Lord had respect unto Abel and to his offering." But when Cain, a tiller of the soil, made an offering of his vegetable produce, the Lord God "onto Cain and to his offering he had not respect."
Due to the social and even religious and spiritual importance of meat, a successful hunter is looked upon as a very special and powerful individual who has great prestige among his band. Moreover, the successful hunter who gives out shares of meat in accordance with his own prerogatives thus gains power over the group as well as respect. In consequence, the successful hunter is sometimes rewarded with more than one wife (or has numerous extra-marital affairs) presumably because he can provide for more than one and by his great stature is actually sought by females as well as prospective father-in-laws
Of course, those who were the best hunters were no doubt the most intelligent of their times, at least insofar as survival is concerned. Supposing that the most intelligent of the race were more likely to breed, with each passing generation, there was a manifold increase in the numbers of intelligent people. That is, the best hunters would have the most wives and thus the most children, whereas those who were not proficient either died out or bred so sparsely that their descendants and genetic contributions simple ceased.
Moreover, females then as now, tend to be attracted to males who hold a more dominant status than they themselves. Males tend to establish relations with females who are subordinate. Hence, our ancient spear throwers, like modern athletes today, often had their pick of willing females thus increasing further their particular genetic contribution to the race.
As noted, however, the ancient female was quite a prized possession, due to her sexual availability and gathering skills. Hence, males had to also compete amongst each other for their favors and attention, particularly in regard to females who may have been similarly dominant and intelligent.
Males also must often fight and sometimes even kill in order to maintain their dominant position, or in order to escape a position of inferiority. Females are not subject to these same pressures, particularly in that males fight not only for dominance but sometimes for sexual access to females which in turn can be a great source of status. These beahviors are most obviously evident among non-human male mammals.
Hence, not only successful hunting, but general intelligence, and the ability to successfully compete against other males in regard to status, dominance, and access to women may have been a tremendous concern of the human male for several thousand years. One might suppose his brain adapted accordingly.
Does this explain, at least in part, the tendency for modern men and adolescent males to sometimes denigrate, even in a teasing manner, the intellectual, physical and financial abilities or even sexual prowess of one other? Is this the remnants of an age old means of achieving dominance?
LANGUAGE & TOOL MAKING, GATHERING, & SHOPPING
The basic skills necessary in the gathering of vegetables, fruits, seeds, berries and the digging of roots include the ability to engage in fine and rapid, temporal-sequential physical maneuvers with the arms, hands, and particularly the fingers. As gathering has been a dominant activity for such a long time period in our prehistory, it is thus not surprising that the brain has possessed rudimentary temporal-sequential capabilities for several million years.
To aid the women on their daily shopping trips they carried large purses (or pouches) made either of leather or the stomach or bladders of various animals into which they could deposit their goods. It probably hung over the woman's shoulder almost like a long strapped purse. Women have thus been gathering (shopping?) and carrying purses for at least 100,000 and perhaps as long as 500,000 years and longer.
The purse was also highly beneficial to the development of social relations. If they did not collect they would probably simply eat what they found on the spot and would have nothing to bring home. In fact, humans are the only primate that engages in gathering. All other primates eat their vegetables as they find them. Of course a few highly social animals such as wolves and wild dogs bring food back to the den in their stomach where they may then regurgitate it for the young, or even for the old, injured or feeble. Birds also bring food to their young.
In addition to gathering, women made tremendous use of tools and may have been the first tool makers. Tool making, like gathering was possibly a major aspect of their lives and one they may have spent considerable time engaged in. In fact, the first tools were made not for hunting but for gathering and rooting plants. That is, if we suppose that gathering predated hunting and scavenging. For example, in grubbing for roots and bulbs, the gatherer would need a digging stick which they probably had to periodically sharpen as they gathered by using stone flakes. They would also carry a hammerstone for cracking nuts and for grinding the various produce collected during the day.
In addition, as the female also would sometimes hunt and try to capture small game (probably with the help of a big friendly dog) she may have carried an ax and knife for the purposes of dispatching whatever hapless prey she chanced upon. By Cro-Magnon times, many of them also carried large flat bone knives up to 9 inches in length.

These ancient women did not spend their time solely gathering, for foods had to be prepared and clothes had to be fashioned out of hides. Her duties may have included cleaning the hides via the use of a scraper, drying and curing the skin over the smoke of a fire, and then using a knife or cutter to make the general desired shape, and then a punch to make holes through which leather straps or vine can be passed so as to create a garment that could keep out the cold. They were also weaving and by Cro-Magnon times they were using a needle to sew garments together. Although there is no way of knowing if woman was the first tool maker, she certainly regularly engaged in these endeavors as a more or less on going activity.
Be it woman or man or both, they eventually developed the capacity to look at and feel a stone or bone and immediately realize its potential as a weapon, tool, or object of art. In this regard, they may have been more attuned to details and had to know stones and bones and the properties and grains of wood, the way, manner and direction it had to be struck and with what instrument to use in order to arrive at the desired effect.
However, in tool making, technique comes to have pre-emmenence. With the invention of technique, a linear, temporal and sequential approach long developed over eons of gathering, probably came to characterize the process. Certain tools are made a certain way with certain instruments with certain movements, and with a certain degree of muscular power and considerable precision. To make and utilize tools requiring a precision grip required that the manufacturer not only have a hand capable of such feats, but a brain that could control this hand and which could use foresight and planning in order to carry out the manufacture.
Due to selective pressures and the survival and breeding of those who were successful at these activities, the left half of the brain, which controls the right hand, became increasingly adapted for the control of temporal-sequencing be it for the purposes of tool making or for gathering. Hence, it was not only tools, but the neural substrate for the temporal-sequential and grammatical aspects of what would become spoken language that was being forged and passed on to succeeding generations, a process that had it's onset several million years ago. As females have engaged in gathering for time periods much longer than males, coupled with her possible role in tool manufacturing and tool use (e.g. skinning, clothes making, etc.), it might be assumed that these changes were maximal in the brains of women, particularly in the motor areas controlling speech (Broca's area) and hand control--and this in turn would explain woman�s propensity to engage in tasks involving fine motor skills, such as typing, knitting, sewing, and so on.




Not only do modern human females demonstrate superior fine motor skills and an earlier onset of language and are less likely to suffer language disturbances, but that it may have been our female ancestors who more fully developed the temporal-sequential aspects of language first as well. Perhaps this was the fruit of knowledge woman offered man only after having taken the first bite. In fact, we find in Genesis that the first gatherer, the first seeker of knowledge, and the first individual to hold a complete conversation (other than God and the serpent) was a woman.
When we consider the many selective pressures which acted on males so as to inhibit speech during the hunt, whereas females were allowed to talk quite freely while gathering (as there would be no fear of scaring off game), it certainly appears that these factors strongly and differentially promoted female linguistic development and a greater capacity to discuss topics unrelated to events associated with the hunt. Language was now being used for social bonding and only later became an instrument designed more and more, at least insofar as males are concerned, for the purposes of exchanging information related to business and sports; i.e. the hunt.
LANGUAGE & CONSCIOUSNESS
Consciousness is not synonymous with language. There are multiple streams of consciousness and mental activity which are subserved by distinct regions of the brain, some of which do not require or even perceive the words of language. It is apparent that other species are conscious and do not require language. And it is obvious other species utilize sensory modalities which are not well developed (olfaction) or non-existant in humans (echo-location), but which make up a great part of the consciousness of these non-human animals. Language is a late evolutionary acquisition, and those aspects of consciousness associated with language, may have been acquired only over the course of the last 35,000 years. Language, and the train of verbal thought, often provide consciousness with contents, and the language-dependent aspects of consciousness are unique to the human mind are are only a recent evolutionary acquisition.
For much of human evolution females have engaged in tasks promoting, requiring, and involving rapid temporal sequential bilateral fine-motor skills such as gathering and domestic-tool construction and manipulation. Given an innate tendency to vocalize more than males and to engage in mutual mother-baby vocalizations, these gathering activities, coupled with prolonged child care and mutual child-mother vocalizations, gave impetus to the evolution of the neocortical speech areas and a female language (and social emotional speech) superiority. The female brain became reorganized and the neocortical representation of older functions were diminished.
In contrast to gathering groups in which females could vocalize with each other and their young, males instead pursued their own violent tendencies, eventually becoming silent hunters of big game. Hunting does not require language, but silence. Over the course of evolution the male proclivity to silently travel long distance in the pursuit of prey exaggerated an already innate male visual-spatial perceptual superiority.
Nevertheless, although hunting does not promote the evolution of speech, males also acquired language skills through maternal genetic inheritance and and as he had a mother who would talk to him and teach him language. Thus, like the proverbial Eve, woman the gatherer provided man the hunter with the fruit of linguistic knowledge and what would become grammatically complex, vocabulary-rich speech, language, and linguistic consciousness.






