






Rhawn Gabriel Joseph, Ph.D.
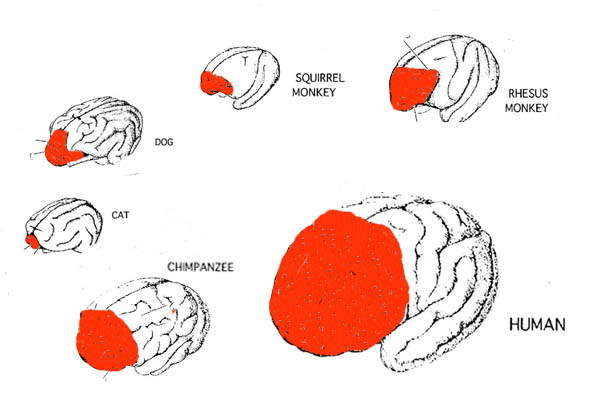
The Evolution of the Frontal Lobes
The frontal lobes serve as the "Senior Executive" of the brain and personality, acting to process, integrate, inhibit, assimilate, and remember perceptions and impulses received from the limbic system, striatum, temporal lobes, and neocortical sensory receiving areas (Fuster 2007; Joseph 2007a; Koechlin et al., 2011; Milner and Petrides 1984; Passingham 1993; Selemon et al. 2013; Shallice and Burgess 1991; Stuss 1992; Stuss and Benson 2007; Strub and Black 1993; Van Hosen et al., 2006).
Moreover, through the assimilation and fusion of perceptual, volitional, cognitive, and emotional processes, the frontal lobes engages in decision making and goal formation, modulates and shapes character and personality and directs attention, maintains concentration, and participates in information storage and memory retrieval (Dolan et al., 2007; Joseph, 2007a, 1988a, 2011a; Kapur et al., 2013; Passingham, 2007; Posner & Raichle, 2014; Tulving et al., 2014).
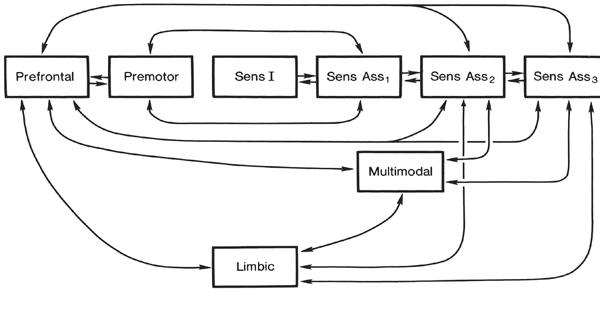
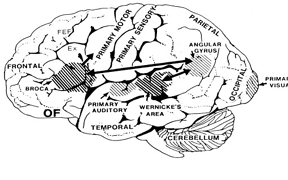
The frontal neocortex is "interlocked" with the limbic system, striatum, and the primary and secondary receiving areas via converging and reciprocal connections, and receives verbal and ideational impulses transmitted from the multi-modality associational areas including Wernicke's area and the inferior parietal lobule (Cavada 1984; Fuster 2007; Jones and Powell 1970; Goldman-Rakic 2013, 2006; Passingham, 1993, 2007; Pandya & Yeterian 1990; Petrides & Pandya 1988). It is thus able to act at all levels of information analysis.
Because of its widespread functional capabilities and interconnections, if damaged there can result excessive or diminished cortical and behavioral arousal, disintegration of personality and emotional functioning, difficulty planning or initiating activity, abnormal attention and ability to concentrate, severe apathy or euphoria, disinhibition and a reduced ability to monitor and control one's thoughts, speech, and actions, including loss of memory.
Moreover, frontal lobe abnormalities commonly result in major cognitive, perceptual, and emotional disturbances, such as schizophrenia, catatonia, mania, depression, obsessive compulsions, aphasia, confabulatory delusions, and the "frontal lobe personality."
Patients may also develop paralysis of the extremeties, or demonstrate severe unilateral neglect of visual-auditory space, or conversely, compulsively utilize tools or other objects, such that in the extreme the right or left hand may act completely independently of the "conscious" mind.


The reason for such a wide range of potential disturbance is that rather than a single pair of frontal lobes there are several "frontal" regions which differ in regard to embryology, phylogeny, cellular composition, functional specificity and interconnection and interactions with other brain areas. The frontal lobes are not a homologous tissue, and each frontal region is concerned with somewhat different as well as overlapping functions (Cummings 1993; Fuster 2007; Goldman-Rakic 2013, 2006; Joseph 2011a; Lura 1980; Pandya and Yeterian 1990; Passingham 1993, 2007; Petrides and Pandya 1988; Selemon et al. 2013; Strub & Black 1993; Van Hoesen, et al., 2006).
Moreover, damage is seldomly restricted to one specific frontal area but commonly disrupts adjoining "frontal" tissues as well. Hence, well localized, as well as a wide ranging spectrum of divergent neuropsychiatric, clinical abnormalities may be induced, sometimes simultaneously, depending on the nature, extent, laterality, and location of the disturbance.
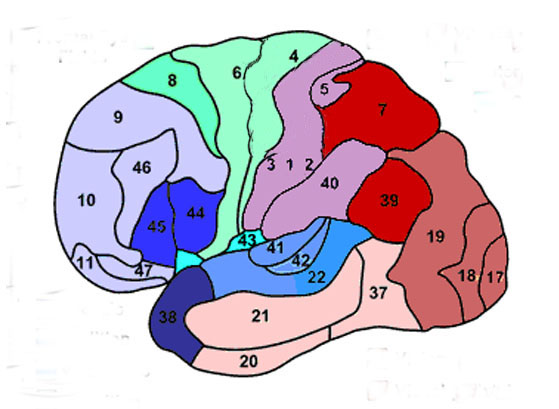
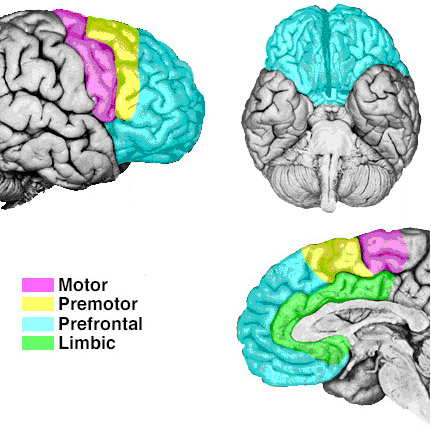
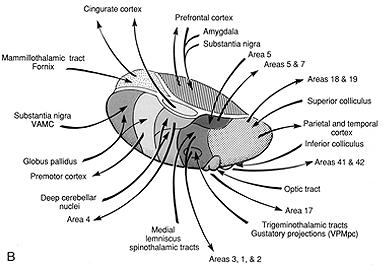
Broadly speaking, the frontal lobes can be functionally lateralized as well as subdivided into four major functional-anatomical regions. As subzones within the right and left frontal lobes, these include the posterior-lateral motor areas (Brodmann's areas 4, 6, 8, 44, 45), the superior lateral convexity (areas 9,8,6,47,46,45,44), the inferior/orbital (areas 47,44,10,11, 12, 13, 14, 25), and the medial motor areas (areas 11,12,10,9,8, 6,32,33,25,24) which overlaps with the motor, lateral convexity, and orbital areas. In addition, the anterior cingulate, and the head of the corpus striatum are intimately associated with and can be considered as part of the "extended frontal lobe."
FUNCTIONAL OVERVIEW:
The right and left frontal lobes appear to exert differing influences over arousal, attention, sexual, emotional, and memory functioning (Brewer, et al., 1998; Joseph, 2007a, 1988a, 2011a; Konishi et al., 2011; Wagner et al., 1998) including even humor appreciation (Shammi & Stuss, 2011). For example, when adults who have been traumatized recall traumatic imagery of when they view combat-related photographs, the right frontal lobe displays increased activity, and the left inferior-orbital region displays reduced activity--as indicating by functional imaging and studies of cerebral blood floow (Rauch et al., 2006; Shin et al., 2007; 2011).

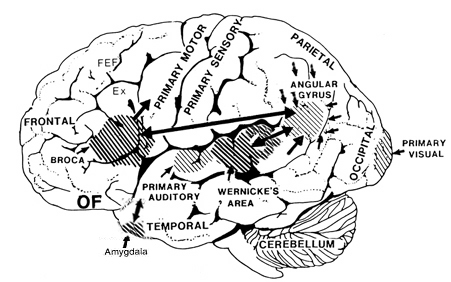
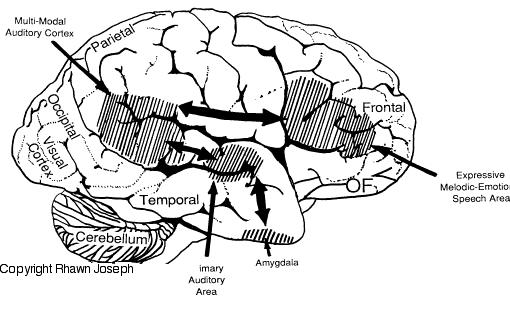
In addition, the right and left frontal lobes are lateralized in regard to language processing, with the right frontal area mediating the expression of emotional and melodic speech (Joseph, 1982, 1988a, 2011a; Ross, 1981, 1993). As is well known, the left frontal area mediates the syntactical, lexical, semantic, and temporal sequential aspects of speech. Because the right and left frontal lobe mediate different aspects of expressive speech, both areas become activated, as indicated by functional imaging, when speaking (Peterson et al., 1988). However, the left frontal lobe is clearly dominant in this regard, as is also demonstrated by functional imaging (Buchel et al., 1998; Demb et al., 2013; Demonet, et al., 2014; Gabrielli et al., 2006; Paulesu, et al., 1993; Peterson et al., 1989; Price, 2007).
On the other hand, although the left frontal lobe is dominant for speech, the right frontal lobe is dominant for arousal and appears to exert bilateral inhibitory influences over arousal whereas the influences of the left frontal lobe are more unilateral and excitatory (Joseph, 1988a, 2011a; Konishi et al., 2011; Tucker, 1981; see also Posner & Raichle, 2014). Hence, if the left frontal lobe is injured, cognitive and expressive functions tend to become suppressed and inhibited--a function not only of the injury but right frontal suppressive influences. By contrast, right frontal injuries are more likely to give rise to disinhibitory states, including the so called "frontal lobe personality"--a function in part, of loss of inhibitory influences.
Specifically, reduced speech output, Broca's aphasia, apathy, "blunted" schizophrenia and major depression are often associated with left lateral (and bilateral) frontal injuries (Bench et al., 2013; Benes, McSparren, and Bird, 1991; Buchsbaum et al. 1998; Carpenter et al. 1993; Casanova et al. 1992; Curtis et al. 1998; d'Elia and Perris 1973, Goodglass & Kaplan, 2011; Hillbom 1951; Perris 1974; Robinson and Downhill 2013; Sarno, 1998). Adults with PTSD also display reduced left frontal lobe activity (Rauch et al., 2006; Shin et al., 2007), whereas sadness can reduce right and bi-frontal activity (Mayberg et al., 2011), though in most instances depression is directly attributed to left frontal dysfunction (see below) and reduced left frontal activity as demonstrated through functional imaging (Bench et al., 2013). As per schizophrenia, it is noteworthy that lateral frontal gray matter reductions and decreased brain volume and activity have been repeatedly noted (Andreasen et al. 1990; Buchanan et al. 1998; Curtis et al. 1998).
By contrast, impulsiveness, confabulation, "motor mouth," grandiosity, and mania are often produced by right frontal (as well as bilateral) lesions (Bogousslavsky et al. 1988; Clark and Davison 1987; Cohen and Niska 1980; Cummings and Mendez 1984; Forrest 1982; Girgis 1971; Jack et al. 2003; Jamieson and Wells 1979; Joseph 2007a, 1988a, 2011a; Lishman 1973; Miller et al. 2007; Oppler 1950; Robinson and Downhill 2013; Rosenbaum and Berry 1975; Starkstein et al. 1987; Stern and Dancy 1942; Stuss and Benson 2007). Moreover, as the right frontal lobe is associated with the expression of emotional melodic speech injuries to this area can also produce pressured and/or confabulatory speech that may be melodically distorted.
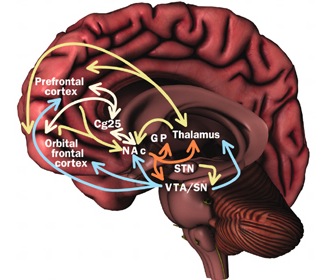
In some respects, orbital frontal dysfunction is similar to right frontal abnormalities in that patients may behave in a labile, impulsive, and emotionally inappropriate fashion. With massive injuries localized to the orbital/inferior areas, and/or which functionally disrupts the frontal-striatal pathways, patients may have difficulty inhibiting unwanted movements or thoughts, and may develop perseverative, obsessive compulsive abnormalities (Baxter et al. 1987; Breier et al. 1992; Buchanan et al. 1993; Iverson and Mishkin 1970; Luria 1980; Malloy, Birhir, and Duffy 1993; Rappoport 1991; Rauch et al. 2014; Swayze et al. 1992). In part, these gross similarities in symptomology are due to the similar role these neocortices play in regard to the control over arousal (e.g. Konishi et al., 2011; Tucker, 1981), the right frontal area regulating neocortical and thalamic activity, the orbital areas exerting inhibitory control over the limbic system (Joseph, 2011a).
By contrast, medial frontal injuries produce symptoms which in some respects are grossly similar to those associated with left frontal injuries, e.g., reduced speech output, mutism, severe apathy. With massive deep medial frontal dysfunction, the patient may become motionless, rigid, and completely catatonic (Hasslet 1980; Laplane et al. 1977; Luria 1980; Penfield & Jasper 1954; Penfield & Welch 1951). However, with less severe injuries, although suffering from disturbances of "will" and volition, patients may also develop "alien hand" symptomology, such that the right or left hand behaves in an independent and "alien" manner--sometimes completely opposite to what the patient intends.
CAVEATS: FUNCTIONAL LOCALIZATION AND SIGNATURE SYMPTOMOLOGY
As detailed below, abnormalities of and damage to the frontal lobes (vs the temporal lobe vs the striatum), can produce signature symptomology. In some cases it is possible to localize a specific subtype of schizophrenia, mania, or depression, to a specific region of the brain based upon those symptoms. Consider, for example, deficits in sacadic eye movements. Smooth pursuit and sacadic eye movements are mediated by the frontal eye fields (Braun, Boman, & Hotson, 2006; MacAvoy et al. 1991) in conjunction with the striatum, midbrain, and cerebellum. Likewise, frontal lobe deficits and reduced ability to perform frontal lobe tests such as the Wisconsin Card Sort, have been found among "Schizophrenics" with smooth pursuit and sacadic abnormalities (O'Discoll, et al. 2013; Rosse et al. 1993), whereas deficits in volitional eye movements have also been documented among those diagnosed with autism (Minshew et al., 2011)--a disorder which shares certain features with schizophrenia.
Of course, localization of pathology should be based on the entire constellation of behavioral and cognitive abnormalities, for collectively they may instead indicate a temporal rather than a frontal lobe injury, or both, and/or widespread reduction in gray matter density, increased ventricular volume, and so on (e.g. Gur et al. 1998; Harrison, 2011; Salisbury et al. 1998; Zipurksy et al. 1998). Unfortunately, psychiatric patients tend to be studied as a group based on a DSM-diagnosis, and with few exceptions (e.g. McGuire et al. 1998) are seldom selected on the basis of similarities in signature symptomology.
Moreover, because the frontal lobes receive converging input from all limbic system structures, the striatum, as well as the primary and association sensory receiving areas, in some instances deficits associated with frontal injury can be produced secondary to abnormalities in these tissues due to disconnection or abnormal activity. This has given rise to considerable confusion and a lack of consensus.
Consider, for example, schizophrenia. Whereas one group of investigators will find evidence of left frontal dysfunction or bilateral gray matter reductions, yet another will report an association with temporal lobe, or ventricular, or striatal abnormalities, whereas yet others will fail to find any clear cut associations at all (reviewed in Harrison, 2011). In consequence, there is little or no consensus as to the neurological foundations of these and other neuropsychiatric disorders, other than the fact that the brain has in some manner been compromised.
Moreover, because the frontal lobes are not a homologous structure but maintain extensive interconnections with a variety of brain regions, sometimes damage to the pathways linking the frontal lobes with these areas, or injuries to these "non-frontal" tissues, can produce similar symptoms.
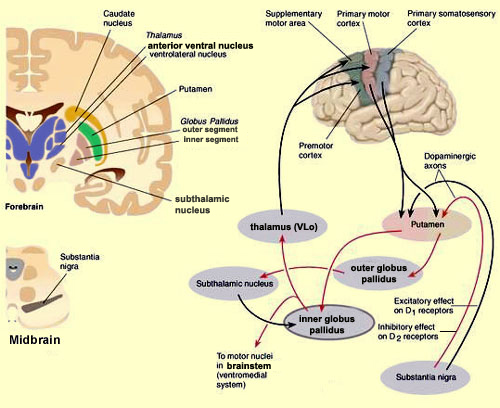
For example, the caudate, putamen, and nucleus accumbens (striatum) are located in the paramedian depths of the right and left frontal lobe, and obsessive-compulsions have frequently been observed not only with frontal injuries, but in association with striatal abnormalities as based on structural (Calabrese, et al. 1993; Robinson, et al. 2013) and functional studies (Perani et al. 2013; McGuire et al. 2014).
Likewise, "schizophrenia" has been associated not only with left frontal injuries, but with left temporal (Bruder et al., 2011; Kwon et al., 2011) or left frontal-temporal lobe injuries (Harrison, 2011), or with damage to the striatum, or the frontal-striatal or temporal (hippocampal/amygdala)-striatal pathways; for example, as measured by positron-emission tomography (e.g. McGuire et al. 1998), P300 evoked potential amplitude (Bruder et al., 2011; Salisbury et al. 1998) and MRI (Jacobsen et al. 1998; Kwon et al., 2011; Nelson et al. 1998; Shidhabuddin et al. 1998), and depending on the diagnostic criteria employed. However, these tissues are functionally linked.
Similarly, mania and confabulatory delusions may be associated with right temporal, right frontal-temporal, and right frontal-parietal dysfunction (Bogousslavsky et al. 1988; Cohen and Niska 1980; Joseph 2007a, 1988a, 2011a; Robinson and Downhill 2013); depending on the diagnostic or research criterion employed as well as the nature of the lesion, e.g. stroke, vs tumor, vs seizure disorder. Again, however, these tissues are linked and often injuries are not always well localized but may compromise adjoining tissues, and even the contralateral frontal lobe--such as due to compression, seizures, etc..
With the caveats in mind, the purpose of this chapter, therefore, is to detail the functional neuroanatomy and to functionally localize and detail the signature symptomology and the clinical psychopathology associated with abnormalities of the different subregions of the frontal lobe and neural pathways including the striatum. Hence, motor functioning, memory, attention, and arousal, schizophrenia, depression, mania, catatonia, obsessive compulsions, aphasia, confabulatory delusions, and the "frontal lobe personality" will be discussed.

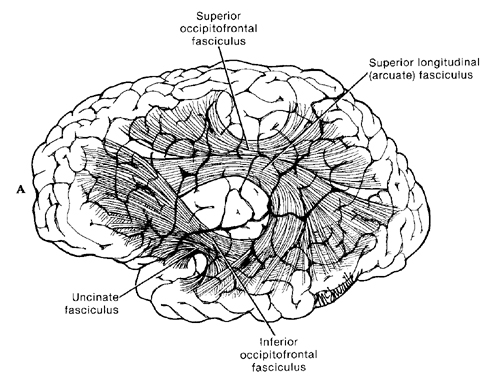
The frontal neocortex is "interlocked" with the limbic system, striatum, and the primary and secondary receiving areas via converging and reciprocal connections, and receives verbal and ideational impulses transmitted from the multi-modality associational areas including Wernicke's area and the inferior parietal lobule (Cavada 1984; Fuster 2007; Jones and Powell 1970; Goldman-Rakic 2013, 2006; Passingham, 1993, 2007; Pandya & Yeterian 1990; Petrides & Pandya 1988). It is thus able to act at all levels of information analysis.
THE FRONTAL LOBE PERSONALITY
With unilateral, bilateral, or even seemingly mild frontal lobe dysfunction patients may initially display an array of waxing and waning abnormalities including the "frontal lobe personality," i.e. tangentiality, childishness, impulsiveness, jocularity, grandiosity, irritability, increased sexuality, and manic excitement (Joseph, 1988a, 2011a; Lishman, 1973). Over fifty years of research and numerous case studies have consistently indicated that with significant frontal lobe pathology attentional functioning may become grossly comprised, behavior may become fragmented, and initiative, goal seeking, concern for consequences, planning skills, fantasy and imagination, and the general attitude toward the future may be lost.
The patient's range of interests may shrink, they may be unable to adapt to new situations or carry out complex, purposive, and goal directed activities, and lack insight, judgement, and common sense (Fuster 2007; Freeman and Watts 1942; Girgis 1971; Hacaen 1964; Joseph 2007a, 1988a, 2011a; Luria 1980; Passingham 1993; Petrie 1952; Stuss 1991; Stuss and Benson 2007). Conversely, when engaging in memory, planning, decision making, goal formation, and tasks requiring imagination, the frontal lobes become highly active--as demonstrated by functional imaging (Brewer et al., 1998; Passingham, 2007; Wagner et al., 1998; Dolan et al., 2007; Squire, et al,. 1992; Tulving et al., 2014; Kapur et al., 2013).
With massive trauma, stroke, neoplasm or surgical destruction (i.e. frontal lobotomy), patients may show a reduction in activity and take very long to achieve very little. They may be unconcerned about their appearance, their disabilities, and demonstrate little or no interest in self-care or the manner in which they dress, or even if their clothes are soiled or inappropriate (Bradford 1950; Broffman 1950; Freeman and Watts 1942; Petrie 1952; Strom-Olsen 1946; Stuss 1991; Stuss and Benson 2007; Tow 1955).
Although some patients demonstrate restlessness, impulsiveness, and flight of ideas, they may also tire easily, show careless work habits and a desire to get things over with quickly. As repeatedly documented following frontal lobotomy, they may immediately develop a tendency to lie in bed unless forcibly removed (Broffman 1950; Freeman and Watts 1942 1943; Rylander 1948; Tow 1955). Even with mild and subtle frontal lobe damage, patients may seem to take hours to get dressed, to finish their business in the bathroom, or to shop and purchase simple items. For example, patients may spend hours in the bathtub playing with the bubbles. A curious mixture of obsessive compulsiveness and passive aggressiveness may be suggested by their behavior.
In severe cases, compulsive utilization of utensils and tools may occur, as well as distractability and perserveration. For example, following frontal lobotomy, "sometimes a pencil and a piece of paper will be enough to start an endless letter that may end up with the mechanical repitition of a certain phrase, line after line and even page after page" (Freeman and Watts 1943, p. 801).
Even with "mild" to moderate frontal lobe injuries patients may initially demonstrate periods of tangentiality, grandiosity, irresponsibility, laziness, hyperexcitability, promiscuity, silliness, childishness, lability, personal untidiness and dirtiness, poor judgment, irritability, fatuous jocularity, and tendencies to spend funds extravagantly. Unconcern about consequences, tactlessness, and changes in sex drive and even hunger and appetite (usually accompanied by weight gain) may occur, coupled with a reduction in the ability to produce original or imaginative thinking.
"Id rather have a bottle in front of me, than a frontal lobotomy" --anonymous
In some respects, damage to the frontal lobes, the right frontal and orbital frontal in particular, can create symptoms similar to a state of alcohol intoxication. This is because alcohol, which is a central nervous system depressant, knocks out and depresses the functioning of the frontal lobes which serve as the senior executive of the brain and personality. If the "senior executive" gets drunk, executive and inhibitory control over the rest of the brain will be impaired.
However, the initial disinhibitory stages of acute intoxication are generally followed by a drunken stupor; the drunk becomes pacified, lethargic, apathetic, and may pay pass out. These symptoms may also result from frontal lobe damage; and some patients may fluctuate between these extremes.



The effects of brain damage on behavior was noted by the ancients and was practiced by physicians even during the middle ages as a means of controlling the behavior of the insane.
Frontal lobotomy, also known as psychosurgery, was born in 1890, precisely because frontal lobe damage can induce apathy. It was in that year that psychiatrist Gottlieb Burckhardt crudely destroyed frontal lobe tissue of six extremely agitated and often violent patients in a psychiatric hospital in Switzerland. Half of these patients became stuporous and were completely pacified. Burckhardt claimed a 50% success rate. However, the other 50% became little different from a wide-awake and violent drunk.
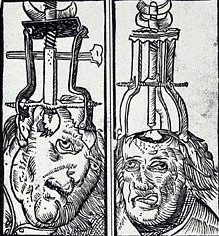
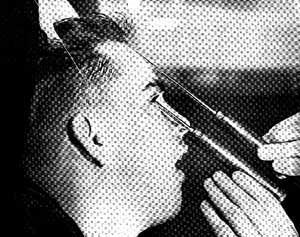
in 1935, a Portuguese physician Ant�nio Egas Moniz drilled holes into the heads of patients and stuck a sharp wire loop into the patient's head and destroyed varying amounts of frontal lobe tissue. He called his surgical technique, "prefrontal leucotomy." Moniz was given the Nobel Prize for medicine in 1949 for this butchery.
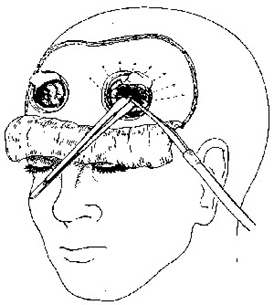
In 1936, Walter Freeman and James Watts developed the surgical ice pick approach, which essentially consisted of sticking a blade into a patient's brain and going "swish swish swish." They called this the "precision method." Most of Freeman's patients were made much worse by the procedure, and Freeman and Watt's took a surprisingly jocular view of these successes:
As stated by Freeman and Watts (1943, p. 805): "Sometimes the wife has to put up with some exaggerated attention on the part of her husband, even at inconvenient times and under circumstances which she may find embarrassing. Refusal, however, has led to one savage beating that we know of, and to an additional separation or two" (p. 805).
Curiously, in these situations Freeman and Watts (1943, p. 805) have suggested that "spirited physical self-defense is probably the best strategy of the woman. Her husband may have regressed to the cave-man level, and she owes it to him to be responsive at the cave-women level. It may not be agreeable at first, but she will soon find it exhilarating if unconventional."
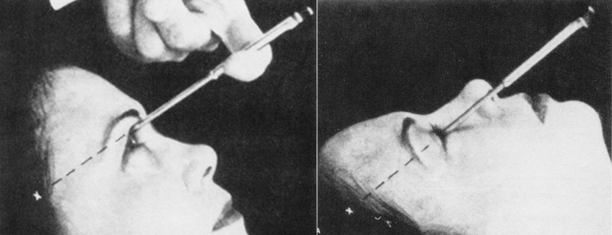
Rosemary Kennedy, the sister of president John F. Kennedy and senator Robert Kennedy, was born mildly retarded. She did not always behave with the class and decorum expected of a Kennedy, and suffered mood swings. Dr. Freeman promised the Kennedy family he could cure her. At age 23, Rosemary was strapped to a table, her head immobilized, and while fully conscious Freeman proceeded to destroy her frontal lobes. He asked her to count and to recite prayers during the procedure and Freeman stopped cutting only after she no longer made any sense. She was "cured" he declared, and in fact, she no longer suffered mood swings. She became little more than a vegetable, unable to speak coherently, or control her bowels, or attend to her most basic needs. She died in a mental institution in 2005.


Following lobotomy or massive or even mild frontal injuries patients may become emotionally labile, irritable, euphoric, aggressive, and quick to anger, and yet be unable to maintain a grudge or a stable mood state as they rapidly oscillate between emotions (Bradford 1950; Greenblatt 1950; Joseph 2007a, 2011a; Rylander 1939; Strom-Olsen 1946; Stuss 1991; Stuss and Benson 2007). Depending on the degree of damage, they may become unrestrained, overtalkative, and tactless, saying whatever "pops into their head", with little or no concern as to the effect their behavior has on others or what personal consequences may result (Broffman 1950; Bogousslavasky et al. 1988; Freeman and Watts 1943; Joseph 2007a, 2011a; Luria 1980; Miller et al. 2007; Partridge 1950; Rylander 1939, 1948; Strom-Olsen 1946).

With severe injuries patients may seem inordinantly disinhibited and influenced by the immediacy of a situation, buying things they cannot afford, lending money when they themselves are in need, and acting and speaking "without thinking." Seeing someone who is obese they may call out in a friendly manner, "Hey, fatty", and comment on their presumed eating habits. If they enter a room and detect a faint odor, its: "Hey, who farted?"
Following severe injuries there may be periods of gross disinhibition which may consist of loud, boisterous, and grandiose speech, singing, yelling, and beating on trays. The destruction of furniture and the tearing of clothes is not uncommon. Some patients may impulsively strike doctors, nurses, or relatives and thus behave in a thoroughly labile, aggressive, callous and irresponsible manner (Benson and Geschwind 1971; Freeman and Watts 1942, 1943; Joseph 2007a, 2011a; Strom-Olsen 1946; Stuss and Benson 2007).
One patient, with a tumor involving the right frontal area, following resection, attempted to throw a fellow patient's radio through the window because he did not like the music. He also loudly sang opera in the halls. Indeed, during the course of his examination he would frequently sing his answers to various questions (Joseph 2007a).
Impulsiveness can also be quite subtle. Luria (1980, p. 294), describes one patient with a slowly growing frontal tumor "whose first manifestation of illness occurred when, on going to the train station, he got into the train which happened to arrive first, although it was going in the opposite direction."

UNCONTROLLED LAUGHTER AND MIRTH
Frontal lobe patients can act in a very childish and puerile manner, laughing at the most trivial of things, making inappropriate jokes, teasing, and engaging total strangers in hillarious conversation (Ackerley 1935; Freeman and Watts 1942, 1943; Kramer 1954; Luria 1980; Petrie 1952; Rylander 1939, 1948; Stuss and Benson 2007). Pathological laughter, joking, and punning may occur superimposed upon a labile effect. Many are vastly amused by their own jokes (Ironside 1956; Kramer 1954; Martin 1950). They can be quite funny, but often they are not!
In part, frontal lobe humor is a function of tangentiality and disinhibition. Loosely connected ideas are strung together in an unusual fashion. The tendency to exaggerate and to impulsively comment upon whatever draws their attention is also contributory, and their humor and laughter may have a contagious quality.
Nevertheless, rather than funny, frontal patients may seem crude and inappropriate. They may laugh without reason and with no accompanying feelings of mirth. These disturbances were in fact documented over 50 years ago. Kramer (1954) for example, describes 4 cases of uncontrollable laughter after lobotomy. They were unable to stop their laughter on command or upon their own volition. The laughter would come on like spells, occurring up to a dozen times a day, and/or continue into the night, requiring sedation in some cases. In these instances, however, the laughter had no contagious aspects but seemed shrill and "frozen." When questioned about the laughter the patients either confabulated a reason for their mirth, or seemed completely perplexed as to the cause.
In general, right orbital damage seems to result in the most severe alterations in mood and emotional functioning (Grafman et al. 2007), which in turn is likely a function of the greater role of the right frontal lobe and the right hemisphere including the right inferior frontal lobe, in the regulation of emotion and arousal (Joseph, 2007a, 1988a, 2011a; Konishi et al., 2011; Tucker, 1981).

In addition to laughter, punning and "Witzelsucht" (puerility), language might become excessively and inappropriately profane, and the patient may seem inordinately inconsiderate, outspoken, and obstinant (Broffman 1950; Partridge 1950; Strom-Olsen 1944; Stuss 1991; Stuss and Benson 2007). However, although they may easily swear, laugh, joke, and make threats, they may also become inordinately apathetic and listless, spending much of their time doing nothing.
Frontal lobe damage and lobotomy reduces one's ability to profit from experience, to anticipate consequences, or to learn from errors (Bianchi 1922; Drewe 1974; Goldstein1944; Halstead 1947; Milner 1964 1971; Nichols and Hunt 1940; Joseph 2007a, 2011a; Petrie 1952, Porteus and Peters 1947; Rylander 1939; Shallice and Burgess 1991; Stuss and Benson 2007; Tow 1955). There is a reduction in creativity, fantasy, dreaming, and abstract reasoning. The capacity to synthesize ideas into concepts or to grasp situations in their entirety is lost, and interests of an intellectual nature are diminished, or sometimes abolished. As described by Freeman and Watts (1943, p. 803) "patients who were great readers of good literature will be interested only in comic books or movie magazines. Men of considerable intellectual achievement... when discussion turns on the great events of the day will pass off some cliches as their own opinions".
In mild or severe cases thinking may be contaminated by perseverative intrusions of irrelevant and tangential ideas, randomly formed associations, and illogical intellectual activity. These patient are also often effected by the immediacy of their environment and have difficulty making plans or adequately meeting long term goals. Even if highly intelligent, they may no longer be able to use that intelligence affectively, and they may undergo a complete personality change.
In 1981, following his graduation from Stanford University, D.F., and three friends, founded their own electronics/computer company, which immediately became a modest success, and began to rapidly grow and expand. D.F., tall, lanky, clumsy, already losing his hair, and only 27 years old, was an electronics genius and in all respects the classic "nerd." He had never dated, and never had a social life, and throughout his younger years had been treated like a "retard" by his school mates. Although he was "vice president" of his company, and a co-owner, and worth over seven figures, he simply lacked the self-confidence to mingle or socialize with the opposite sex, and didn't feel comfortable interacting with anyone who did not share his enthusiasm for computers. Neverthless, D.F. was lonely. He needed a girlfriend.
In 1985 his company "went public" (made a public offering of stock) and D.F. was suddenly a multi-millionaire without a girlfriend. It was soon thereafter that a "new girl" was hired to work as an assistant and secretary, just outside his office. "She was beautiful! An angel! Long blond hair. Green eyes. She must have weighed only 100 pounds. Everytime I walked out side my office I felt shocked to see her. And she wasn't married. She was only 23. I started thinking about her all the time. I coudn't concentrate. I dreamed about her. I wanted to marry her. I loved her. Love her!" he gushed, his words racing.
But D.F. didn't have the nerve to approach her, or talk to her, except to say hi. "She always smiled at me, when I said hi. A big smile. I knew she liked me." But he still didn't ask her out. Instead, he began to fantasize, about being married, having children.
And then after several tortured months, D.F. came to a decision. "I bought her a huge diamond ring. Ten thousand dollars. And I asked her to marry me."
According to D.F., he simply approacher her, offered her the ring, which she took and put on her finger. And then he asked her to marry him. Apparently she was stunned. Apparently she even laughed. Apparently she even thanked him for the ring. But she said no.
D.F. was mortified and became massively depressed. He couldn't function. Couldn't think."I just wanted to die." D.F. purchased a handgun. Took it home. Placed the barrel in his mouth. But then aimed it incorrectly. He pointed it directly into the roof of his mouth, pulled the trigger, and then blew out his right frontal lobe.
Approximately six months later, D.F. arrived for his appointment to see me, with a toothbrush, toothpaste, hair brush, and wash rag sticking out of his shirt pocket (Joseph 1988a). When I asked, pointing at his pocket, "What's all that for?" he replied with a laugh, "That's just in case I want to brush my teeth," and in so saying he quickly drew the toothbrush from his pocket and began to demonstrate. During the course of the exam he laughingly demonstrated how the skin flap which covered the hole in his head (from the craniotomy and bullet wound) could bulge in or out when he held his breath or held his head upside down. He even climbed up and stood on this examiner's desk and bent over so as to offer a better view of the hole in his head.
Throughout the exam he behaved in a silly, puerile manner, often joking and laughing inappropriately. Even when discussing the blond secretary and the aftermath, he laughed, becoming serious for only a few moment when I asked him if she had given him back the ring. She hadn't. Nevertheless, despite his bizarre and inappropriate behavior, D.F.'s overall WAIS-R IQ was above 130 (98% rank: "Very Superior"). Unfortunately, although he had a high IQ, he could no longer employ that intelligence, intelligently. On the other hand, he was no longer depressed, which is one of the many reasons that frontal lobotomies became such a popular in-office psychiatric procedure during the 1940s and 1950s.
Frontal lobe patients also may have difficulty thinking up or considering alternative problem solving strategies and thus developing alternative lines of reasoning. For example, Nichols and Hunt (1940) dealt a patient five cards down including the ace of spades which always fell to the right on two successive deals and then to the left for two trials. The patients task was to learn this pattern and turn up the ace. The patient failed to master this after 200 trials.

Some frontal lobe patients may also have extreme difficulty sorting even common everyday objects according to category (Rylander, 1939; Tow, 1955), for example, sorting and grouping drinking containers (glasses) with other drinking containers (mugs), tools with tools, etc. Similarily, they may have difficulty performing the Wisconsin Card Sorting Task (Crockett et al., 2007; Drewe, 1974; Milner, 1964, 1971) which involves sorting geometric figures according to similarity in color, shape, or number. However, the manner in which a patient fails on this task is dependent on the locus and laterality of the damage.
For example, patients with orbital damage seem to have relatively little difficulty performing this category sorting task (Drewe, 1974; Milner, 1971). Similarly, patients with right frontal damage, although they show a tendency to make perseverative type errors (i.e. persisting in a choice pattern which is clearly indicated as incorrect), perform significantly better than those with left frontal damage (Drewe, 1974; Milner, 1964, 1971). Thus overall, patients with left medial and convexity lesions perform most poorly, and have the greatest degree of difficulty thinking in a flexible manner or developing alternative response strategies.
I.Q. Testing
A number of studies of conceptual functioning have been performed before and after surgical destruction of the frontal lobes. Although in some cases, such as D.F., described above, the IQ remains high, performance is so uneven and there is so much intertest variability that it is apparent that patients have suffered significant declines (Petrie, 1952; Smith, 1966). In studies in which patients undergoing frontal leucotomy for intractable pain were administered the Wechsler Intelligence Scales both pre- and post surgery, a 20 point drop in the IQ was reported (Koskoff, 1948, cited by Tow, 1955).
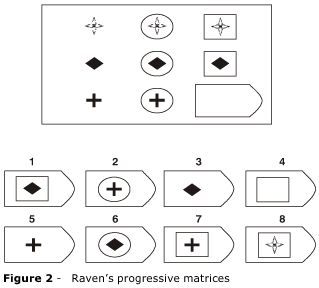
Likewise, in cases where the Raven's Progressive Matrices or Porteus Mazes were administered both before and after lobotomy, significant declines in intellectual functioning have been documented (Petrie, 1952; Porteus & Peters, 1947; Tow, 1955). As with most tests, the usual pattern is to improve with practice. Hence, these results (and those mentioned above) indicate that frontal lobe damage disrupts abstract reasoning skills, verbal-nonverbal pattern analysis, learning and intellectual ability, as well as the capacity to anticipate the consequences of one's actions or to profit from experience. However, the effects of frontal damage on IQ is dependent on the locus of the damage.
For example, left frontal patients show lower Wechsler IQs than those with right frontal lesions (Petrie, 1952; Smith, 1966). In fact, 17 of 18 patients with left frontal damage reported by Smith (1966) scored lower across all subtests compared to those with right frontal lesions. Indeed, patients with left sided destruction perform as poorly as those with bilateral damage (Petrie, 1952).
In analyzing subtest performance, Smith (1966) notes that left frontal lobotomy patients scored particularly poorly on Picture Completion (which requires identification of missing details). This is presumably a consequence of the left cerebral hemisphere being more conerned with the perception of details (or parts, segments) vs wholes (chapters 10, 11). Petrie (1952), however, reports that performance on the Comprehension subtests (i.e. judgment, common sense) was most significantly impaired among left frontals.
In contrast, individuals with severe right frontal damage have difficulty performing Picture Arrangement--often leaving the cards in the same order in which they are laid (McFie & Thompson, 1972). This may be a consequence of deficiencies in the capacity to discern social-emotional nuances, a function at which the right hemisphere excels (chapter 10).
Nevertheless, since so few studies have been conducted it is probably not reasonable to assume that lesions lateralized to the right or left frontal lobe will always effect performance on certain subtests, particularly if there is a mild injury. It is also important to consider in what manner lateralized effects on IQ may be contributing to or secondary to reduced motivation and apathy since bilateral and left frontal damage often give rise to this constellation of symptoms. If the patient is apathetic they are not going to be motivated to perform at the best of their ability.
Thus, where with left frontal injuries patients may seem apathetic, indifferent, and/or severely depressed and psychotic if not schizophrenic, right frontal injuries are associated with manic-like disinhibited states, including waxing and waning abnormalities associated with manic-depression. Patient's may become so disinhibited they develop the classic "frontal lobe personality," and become disinhibited, hyperactive, euphoric, extroverted, labile, overtalkative, and may develop perseveratory tendencies. Patients may become so disinhibited, delusional, grandiose, and emotionally labile that they develop what has classically been described as mania.
By contrast, with a left frontal lesion, rather than a loss of emotional control, there is a loss of emotion, and the patient will become severely apathetic, indifferent, and with massive lesions unresponsive, though classically left frontal injuries are associated with depression.
These right and left frontal differences are a function of lateralized differences in the control over arousal. Whereas the orbital frontal lobes contral limbic arousal, the right and left frontal lobes contraol neocortical arousal, with the right frontal lobe exerting bilateral inhibitory and excitatory influences, whereas the left frontal lobe exerts unilateral excitatory influences.
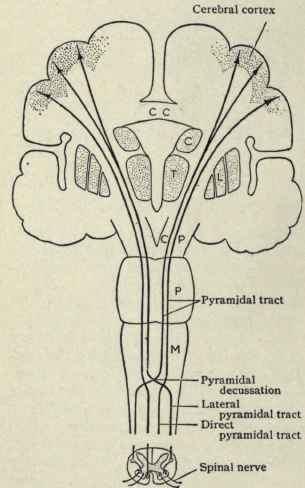
FRONTAL LOBE REGULATION OF THE NEOCORTEX
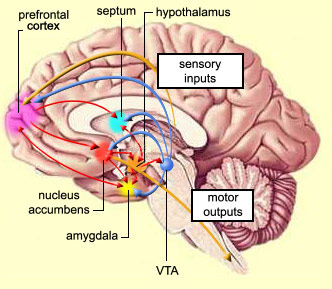
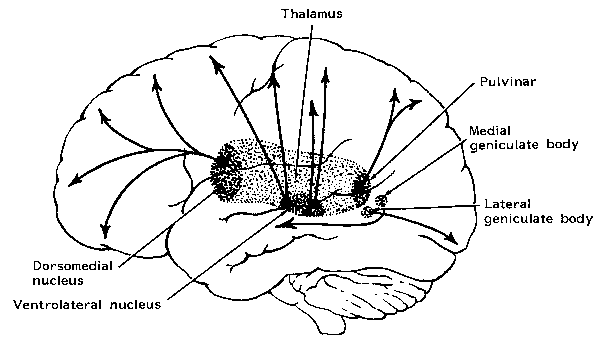
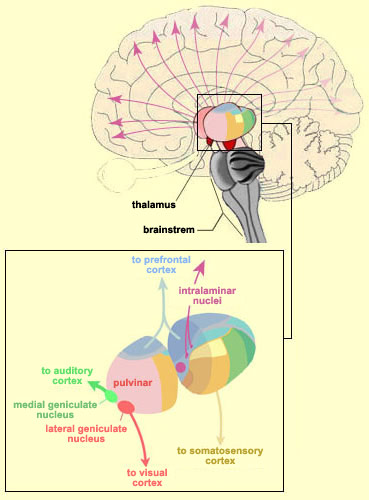
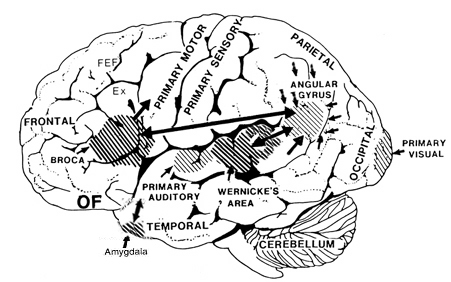
With the exception of olfactory information, which via the olfactory tracts projects to the limbic system and is relayed as well as directly transmitted to the orbital region (Cavada, 1984; Gloor, 2007), all sensory impulses are first transferred to the thalamus before being transmitted to the primary auditory, visual, and somesthetic receiving areas. From the primary zones this information is sent to 3 separate major locations: to the immediately adjacent sensory association area, back to the thalamus, and to the motor cortex of the frontal lobes.

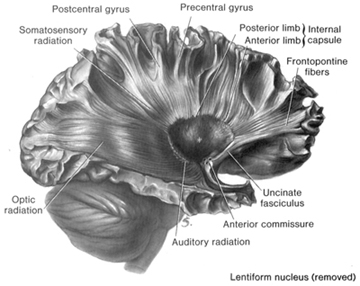
Fibers passing to and from the thalamus and the cortical sensory receiving areas give off collaterals to the reticular thalamic nucleus--which in addition sends fibers which envelop and innervate most of the other thalamic nuclei (Scheibel & Scheibel, 1966; Updyke, 1975). The reticular thalamus maintains reccurent inhibitory interconnections with other thalamic neurons (Huntsman et al., 2011) and acts to synchronize and selectively gate transmission from the thalamus to the neocortex and continually samples thalamic-cortical activity (Skinner & Yingling, 1977; Yingling & Skinner, 1977).
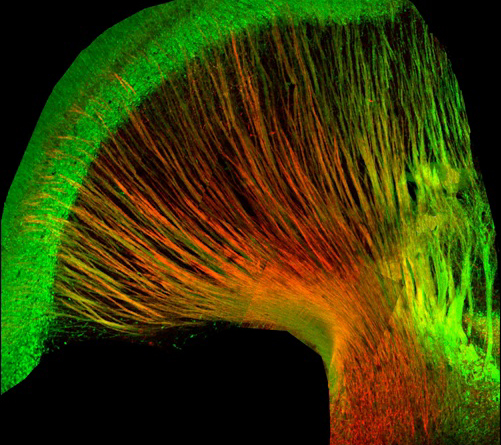
Thalamic Cortical Axons
The reticular thalamus is controlled by the lateral convexity of the frontal lobes, and the lateral portion of the dorsal medial thalamus with which it maintains dense interconnections (Skinner & Yingling, 1977; Yingling & Skinner, 1977). The convexity and lateral dorsal medial nucleus (LDM) are also richly interconnected and together exert significant steering influences on the reticular thalamus. That is, the lateral frontal convexity appears to exert specific influences on the LDM so as to promote or diminish the flow of information to the cortex and thus modulate specific perceptual and cognitive activities occuring within the neocortex --activity which it is simultaneously sampling. This is in contrast to the orbital region with its connections to the reticular formation and the medial magnocellular segment of the dorsal medial thalamus, and its influences on generalized arousal and limbic activation/inhibition.
To recapitulate, the lateral frontal system is able to influence cognitive/perceptual cortical functioning via the sampling of activity occurring throughout the neocortex at all levels of informational analysis, and via its modulating influences on the lateral portion of the dorsal medial and reticular thalamic nuclei. The lateral frontal region is thus able to act at any stage of processing, from initial reception to motor expression so as to facilitate or inhibit further analysis, selectively acting to determine exactly what type of processing occurs throughut the neocortex.
Via integration and inhibitory action and through its neocortical and thalamic links the lateral convexity it is able to coordinate interactions between various regions of the neuroaxis so as to organize, mobilize, and direct overall cortical and behavioral activity and to minimize conflicting demands, impulses, distractions and/or the processing of irrelevant information.
When damaged, depending on the site (e.g. inferior vs superior convexity) or laterality of the lesion, there can result behavioral disinhibition, flooding of the sensory association areas with irrelevant information, hyperreactively, distractability, memory loss, impulsiveness, and/or apathy, reduced motor-expressive activities (e.g. speech arrest), and sensory neglect (Como et al. 1979; Fuster, 2007; Joseph 2007a, 2011a, Joseph et al. 1981; Passingham, 1993). Similar disturbances can result when the dorsal medial nucleus or the bi-directional pathways linking the thalamus and frontal lobe are severed (Graff-Radford et al.1990; Skinner & Yingling, 1977; Victor et al. 1989).
Hence, in summary, the oribtal region exerts modulating influences on subcortical and geralized limbic arousal. By contrast, the lateral convexity of the frontal lobes are "interlocked" with the the sensory receiving areas (Cavada 1984; Fuster 2007; Jones et al. 1978; Jones and Powell 1970; Joseph, 2011a; Pandya and Kuypers 1969) and maintains rich interconnections with the reticular and dorsal medial nucleus of the thalamus (Skinner and Yingling 1977; Yingling and Skinner 1977) which relays sensory impressions to the neocortex.
The lateral frontal lobes, therefore are able to sample perceptual input as it is received in the thalamus, and thus prior to and after it has been transferred to the neocortical receiving areas. Through its interconnections with the primary and association areas, the frontal lobes can also censor, inhibit, and thus control the processing of this data, and in this manner can control attention as well as facilitate or inhibit further analysis and thus information processing throughout the neocortex. These frontal capabilities include information storage and retrieval at the neocortical level; i.e. memory (Brewer et al., 1998; Carpenter et al., 2011; Hasegawa et al., 1998; Koechlin et al., 2011; Wagner et al., 1998).
In consequence, when the lateral regions are injured, selective attention and memory functioning may become impaired such that individuals may experience sensory-perceptual overload, become distractible, disinhibited, confused, and have difficulty keeping their mind on a certain tasks and/or recalling and acting upon events planned for the future. Similar disturbances can result when the dorsal medial nucleus or the bi-directional pathways linking the thalamus and frontal lobe are severed (Graff-Radford et al.1990; Skinner and Yingling 1977; Victor et al. 1989). However, the nature of these disturbances depends on if the right vs left frontal lobe has been negatively impacted.
Whereas left frontal injuries result in reduced functional and expressive activity, right frontal injuries are more likely to be associated with disinhibition, hyperreactively, distractability, impulsiveness, and the flooding of the sensory association areas with irrelevant information (Joseph 2007a, 1988a, 2011a). This is because the right frontal lobe is dominant over the left, and exerts bilateral influences in the regulation of attention and arousal (Cabeza and Nyberg 2007; DeRenzi and Faglioni 1965; Dimond 1976 1979; Heilman and Van Den Abell 1979 1980; Jeeves and Dixon 1970; Joseph 2007a, 1988ab, 2011a; Konishi et al., 2011; Pardo et al. 1991; Tucker 1981). Thus, if the left frontal lobe is damaged, the right, now acting unopposed may exert bilateral inhibitory influences and thus hypo-arousal.
For example, the intact, normal right hemisphere is quicker to react to external stimuli, and has a greater attentional capacity compared to the left (Dimond, 1976, 1979; Heilman & Van Den Abell, 1979; Jeeves & Dixon, 1970; Joseph, 1988ab). In split brain studies the isolated left hemisphere tends to become occasionally unresponsive, suffers lapses of attention, and is more limited in attentional capacity as compared to the right which attends to both the left and right half of visual, auditory, and tactile space (Dimond 1976, 1979; Joseph 2007a, 1988ab). Indeed, visual and somesthetic stimuli, or active touch exploration with either the right or left hand, elicits evoked EEG responses preferentially and of greater magnitude over the right hemisphere and right frontal lobe (Desmedt 1977).
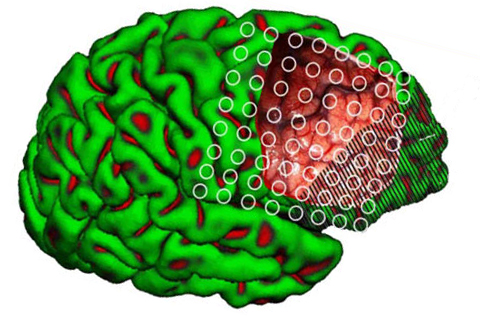
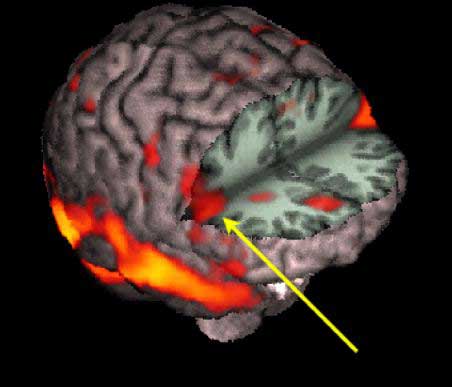
The right hemisphere also becomes desynchronized (aroused) following left or right sided stimulation (indicating it is bilaterally responsive), whereas the left brain is activated only with unilateral (right side) stimulation (Heilman & Van Dell Abell, 1980). In fact, the right frontal lobe and hemisphere responds more quickly even to stimuli appearing on the right side (Dimond 1976 1979; Heilman and Van Den Abell 1979, 1980; Jeeves and Dixon 1970). In consequence, if the right frontal lobe (or hemisphere) is injured, the left frontal lobe is unable to attend to events occurring on the left side of the body, which results in unilateral left-side neglect.
The right frontal lobe is also larger than the left suggesting a greater degree of interconnections with other brain tissue, and it appears to exert bilateral inhibitory influences on attention and arousal (Cabeza and Nyberg 2007; DeRenzi and Faglioni 1965; Dimond 1976 1979; Heilman and Van Den Abell 1979, 1980; Jeeves and Dixon 1970; Joseph 2007a, 1988ab; Konishi et al., 2011; Pardo et al. 1991; Tucker 1981). For example, as based on functional imaging, it has been found that when performing a "go/no-go task" and the Wisconsin Card Sort--tasks requiring the inhibition of irrelevant or erroneous responses, activity significantly increases in the right inferior frontal lobe, and that this was the case irrespective of if the human subjects used the right or left hand (Konishi et al., 2011). By contrast, the left frontal region appears to exert unilateral excitatory influences which promotes right sided motor control and speech expression.
However, because the right frontal lobe appears to exert bilateral inhibitory influences, whereas the influences of the left are unilateral and excitatory, when the left frontal region is damaged, the right may act unopposed and there may be excessive left cerebral inhibition or reduced activity (e.g., Bench et al., 2013); for example, as manifested by speech arrest, depression, and/or apathy.
Nevertheless, because left cerebral excitatory influences are predominantly unilateral, with massive right cerebral damage, although the left hemisphere is aroused, the left is unable to activate the right half of the brain. This may result in unilateral inattention and neglect of the left half of the body and space (Heilman & Valenstein, 1972; Joseph, 2007a, 1988a; Na et al., 2011). That is, the patient's (undamaged) left hemisphere may ignore his of her left arm or leg, and if their neglected extremities are shown to them, may claim they belong to the doctor or a person in the next room. That such disturbances occur only rarely with left frontal or left hemisphere damage further suggests that the right hemisphere is able to continue to monitor events ocurring on either side of the body. Thus although the damaged left hemishere is hypoaroused (or inhibited by the right), there is little or no neglect.
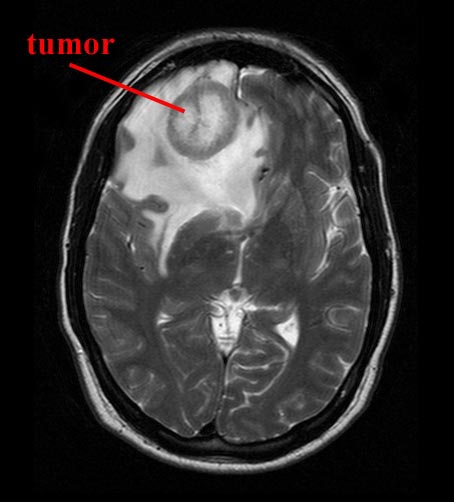
However, with lesions involving the right frontal lobe, not only is there a loss of inhibitory control, but the left may act unopposed such that there is excessive excitement. The patient becomes disinhibited as manifested by speech release, confabulation, lability, and a host of impulsive disturbances which may wax and wane in severity.
As detailed below, mania, confabulation, hypersexuality, tagentiality, and impulsive, labile, disinhibited and inappropriate social and emotional behaviors are predominately associated with right frontal dysfunction (Bogousslavsky et al. 1988; Clark and Davison 1987; Cohen and Niska 1980; Cummings and Mendez 1984; Fischer et al. 2013; Forrest 1982; Girgis 1971; Jack et al. 2003; Jamieson and Wells 1979; Joseph 2007a, 1988a, 2011a; Kapur and Coughlan 1980; Lishman 1973; Miller et al. 2007; Oppler 1950; Rosenbaum and Berry 1975; Shapiro et al.1981; Starkstein et al. 1987; Stern and Dancy 1942; Stuss and Benson 2007); a function of loss of inhibitory control as well as its role in all aspects of emotion. In fact, and as originally pointed out by Lisman (1968, 1973), injuries to the right frontal lobe are clearly associated with what has been described as the "frontal lobe personality," including, in the extreme, the development of a full blown manic psychosis coupled with disinhibited sexuality.

DISINHIBITED SEXUALITY
In some cases following frontal lobe damage patients may engage in inappropriate sexual activity (Benson and Geschwind 1971; Brutkowski 1965; Freeman and Watts 1942, 1943; Girgis 1971; Leutmezer et al., 2011; Lishman 1973; Miller et al. 2007; Strom-Olsen 1946; Stuss and Benson 2007). One patient, after a right frontal injury began patronizing up to 4 prostitutes a day, whereas his premorbid sexual activity had been limited to Tuesday evenings with his wife of 20 years (Joseph 1988a).

Another patient with a right frontal stroke propositioned nurses and would spontaneously reach out and fondle large breasted women (Joseph 1988a).
It is not unusual for a hypersexual, disinhibited frontal lobe injured individual to employ force. One individual who was described as quite gentle and sensitive prior to his injury, subsequently raped and brutalized several women. Similar behavior has been described following lobotomy.

Seizure activity arising from the deep frontal regions have also been associated with increased sexual behavior, including sexual automatisms, exhibitionism, gential manipulation, and masturbation (Leutmezer et al., 2011; Spencer, Spencer, Williamson, & Mattson 2003; Williamson, et al., 1985). One young man that I evaluated and who was subsequently found, with depth electrode recording, to have seizures emitting from the right frontal lobe, had been arrested over 7 times for exposing himself in public. His parents complained that he would sometimes walk around the house grabbing and exposing his genitals, and would sometimes even pee on the floor. In fact, while I was evaluating him as he lay in bed at the Yale Seizure Unit (VAMC) he suffered a seizure which involved the following sequence. He grunted loudly and his left arm shot out in a lateral arc. His left hand then returned to his body and he began to fiddle with the buttons of his pajamas continuing in a downward motion until he reached his penis which he then took in his hand and began to squeeze. As I looked on, he suddenly began to urinate and with such force that I was nearly sprayed with urine. Fortunately, I deftly escaped by leaping to the side and against the wall which put me well out of his range.
By contrast, a young woman I examined with right frontal-temporal seizures would spread her legs and engage in pelvic thrusting, coupled with grunting, lip licking and tongue protrusion. Currier et al., (1971), have also reported pelvic thrusting and moaning, and sex appropriate vocalizations, with temporal lobe seizures.
However, according to Leutmezer et al., (2011) and as based on prolonged scalp-EEG monitoring, sexual automatisms, such as "sexual hypermotoric pelvic or truncal movements are common in frontal lobe seizures," whereas "discrete genital automatisms, like fondling and grabbing the genitals are more common in seizures involving the temporal lobe." Presumably, the results of Leutmezer et al., (2011) differ from that of Currier, et al., (1971), Joseph (1988a, 2011a), Spencer et al., (2003), and Williams et al., (1985), due to their use of scalp rather depth electrodes which are more sensitive and exacting. On the other hand, temporal lobe/hippocampal sclerosis and atrophy were also documented in the Leutmezer et al. (2011) study.
Nevertheless, given the close functional association between the frontal and temporal lobes, and the fact that even a frontal seizure can propagate to the amygdala and thus involve the temporal lobe, perhaps the dysfunctional differences in abnormal sexual behavior are due to seizure origin and the subsequent spread of seizure activity.

There is also some evidence for functional laterality in regard to sexual automatisms and abnormal sexual behavior. In most instances, "sexual" seizures are associated with right frontal seizure foci (Joseph 1988a; Spencer et al. 2003). However, patients may also become hyposexual (Greenblatt 1950; Miller et al. 2007), especially with left frontal injuries, and/or experience genital pain with left temporal seizures (Leutmezer et al., 2011).
Mania and manic-like features have been reported in many patients with injuries, tumors, and even seizures involving predominantly the frontal lobe and/or the right hemisphere (Bogousslavsky et al. 1988; Clark and Davison 1987; Cohen and Niska 1980; Cummings and Mendez 1984; Forrest 1982; Girgis 1971; Jack et al. 2003; Jamieson and Wells 1979; Joseph 2007a, 1988a, 2011a; Lishman 1973; Miller et al. 2007; Oppler 1950; Robinson and Downhill 2013; Rosenbaum and Berry 1975; Starkstein et al. 1987; Stern and Dancy 1942).
One frontal patient described as formerly very stable, and a happily married family man, became excessively talkative, restless, grossly disinhibited, sexually preoccupied, extravagantly spent money and recklessly purchased a business which soon went bankrupt (Lishman 1973).
In another case, a 46-year old woman was admitted to the hospital and observed to be careless about her person and room, and incontinent of urine and feces. She slept very little and acted in a hypersexual manner. Her symptoms had developed several months earlier when she began accusing a neighbor of taking things she had misplaced. She also would confront him and strip off her clothes. She began going about in just a slip and bra, and informed people she was descended from queens, was fabulously wealthy, and that many men wanted to divorce their wives and marry her. During her hospitalization she was frequently quite loud, disoriented to time and place, and extremely tangential, jumping from subject to subject. After several years she died and a meningioma involving the orbital surface of the right frontal lobes was discovered (Girgis 1971).
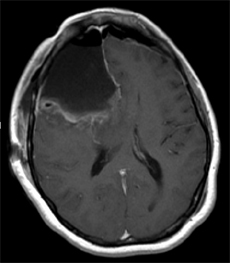


I have examined 19 male and five female patients who developed mania after suffering a right frontal stroke or trauma to the right frontal lobe. All but four of the males had good premorbid histories and had worked steadily at the same job for over 3-5 years (e.g. Joseph, 2007a, 1988a). Following their injuries all developed delusions of grandeur, pressured speech, flight of ideas, decreased need for sleep, indiscriminant financial activity or irresponsibility, emotional lability, and increased libido, including, in one case, persistent sexual overtures coupled with genital exposure, to the patient's sisters and mother.
One formerly very conservative engineer with over 20 patents to his name suffered a right frontal injury when he fell from a ladder. He became sexually indiscriminate and reportedly patronized up to 3 prostitutes a day, whereas before his injury his sexual activity was limited to once weekly with his wife. He also spent money lavishly, suffered delusions of grandeur, camped out at Disney Land and attempted to convince personnel to fund his ideas for a theme park on top of a mountain, and at night had dreams where the Kennedy's would appear and offer him advice --and he was a republican!
Although language is usually discussed in regard to grammar and vocabulary, it is also emotional, melodic and prosodic --features which enable a speaker to convey and a listener determine, intent, attitude, feeling and meaning (chapters 10, 15). A listener comprehends noy only what is said, but how it is said--what a speaker feels.
Feeling and attitude are conveyed through the melody (musical qualities), inflection, intonation, and prosody of one's voice, and by varying the pitch, inflection, timbre, stress contours, melody, as well as the rate and amplitude of speech --capacities predominantly mediated by the right half of the cerebrum (see chapter 10).
Patients with severe forms of Broca's expressive aphasia are unable to discourse fluently. However, they may be capable of swearing, making statements of self-pity, praying, singing, and even learning new songs (Gardner 1975; Goldstein 1942; Gorelick and Ross 1987; Joseph 1988a; Ross 1981; Smith 1966; Smith and Burklund 1966; Yamadori, Osumi, Mashuara, and Okuto, 1977)--although in the absence of music they would be unable to say the very words they had just sung. This is because the ability to produce non-linguistic and musical/emotional sounds is mediated by the undamaged right frontal lobe and limbic nuclei (Gardner 1975; Gorelick and Ross 1987; Joseph 1988a; Ross 1993; Shapiro and Danly 1985). Indeed, just as the left frontal convexity (i.e. Broca's area) subserves the syntactical, temporal-sequential, motoric, and grammatical aspects of linguistic expression (Foerster 1936; Fox 2013; Goodglass & Kaplan, 2000; LeBlanc 1992; Petersen et al. 1988, 1989; Sarno, 1998), there is a homologous region within the right frontal area which mediates the expression of emotional and melodic speech (Gorelick and Ross 1987; Joseph 1982, 1988a, 2011a; Ross 1981, 1993; Shapiro and Danly 1985).
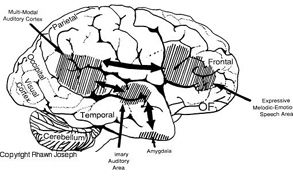
With massive damage involving the right frontal melodic-emotional speech area, speech may become flat and monotonous, or conversely, the ability to alter or convey melodic and prosodic elements may become exceedingly abnormal and distorted. With extensive injuries to the right frontal lobe, patients may lose control over their voice and at times may sound as if they are crying, wailing, or screeching. Such patients may loose the ability to engage in vocal mimicry or to accurately repeat various statements in an emotional manner (Gorelick and Ross 1987; Joseph 1988a; Ross 1981, 1993).
With mild damage, rather than severe distortions or a loss of melody, the intonational qualities of the voice can become mildly abnormal and patients may seem to be speaking with an odd midwestern-like accent--particularly with deep lesions of the right frontal area, perhaps involving the cingulate or basal ganglia. Prosodic distortion in the form of an unusual accent is sometimes seen in seizure disorders involving deep right frontal or frontal-temporal areas.
On the otherhand, with left frontal lesions some patients develop what sounds like an unlearned foreign accent, as if they were from Germany, France, etc. (Blumstein et al. 1987; Graff-Radford et al. 2007). This is due, in part, to distortions involving the pronounciation of vowells.
When damage is limited to this right frontal emotional-motor speech area, the ability to comprehend and understand prosodic-emotional nuances appears to be somewhat intact (Gorelick & Ross, 1987; Joseph, 1988a; Ross, 1993). However, with right temporal injuries, the ability to comprehend these nuances may be lost (Ross, 1993).
It is interesting to note, however, that despite the clinical, neuropsychological, and neuroanatomical evidence indicating that the right frontal and right temporal areas contribute to the production of emotional prosodic melodic speech, that functional imaging and blood flow studies have failed to display significant activity in these regions during language-related activities (Frost, et al., 2011; Pujol, et al., 2011)--a function perhaps, of the insensitivity of the tasks and/or measures employed in this regard.
One frontal patient when asked what he received for Christmas replied, "I got a record player and a sweater." (Looking down at his boots) "I also like boots, westerns, popcorn, peanuts and pretzels." Another right frontal patient when asked in what manner an orange and a banana were alike replied, "fruit. Fruitcakes--ha ha--tooty fruity." When asked how a lion and a dog were alike he responded, "They both like fruit--ha ha. No. That's not right. They like trees--fruit trees. Lions climb trees and dogs chase cats up trees, and they both have a bark."
Tangentiality is in some manner related to impulsiveness as well as circumlocution. In contrast, patients with circumlocutious speech often have disturbances involving the left cerebral hemisphere and frequently suffer from word finding difficulty and sometimes receptive or expressive dysphasia. They experience difficulty expressing a particular idea or describing some need as they have trouble finding the correct words. Thus talk around the central point and only through successive approximations are able to convey what they mean to say.



Patients with tangential speech lose the point altogether. Instead, words or statements trigger other words or statements which are related only in regard to sound (e.g. like a clang association) or some obscure and ever shifting semantic category. Speech may be rushed or pressured and the patient may seem to be free associating as they jump from topic to topic.
Hence, in contrast to the aphasia, or speech arrest associated with left frontal injuries, right frontal lesions may result in speech release ("motor mouth"). Speech becomes disinhibited, pressure, and contaminated with tangential associations, and the patient may seem to be free associating as they jump from topic to topic. In the extreme speech becomes filled with confabulatory ideas.
When secondary to right (or bilateral) frontal damage, speech may become exceedingly bizarre, delusional, and fantastical, as loosely associated ideas become organized and anchored around fragments of current experience.
For example, one patient, when asked as to why he had been hospitalized, denied that there was anything wrong, but instead claimed he was there to do some work. When asked what kind of work, he pointed to the air conditioning unit, and stated: "I'm a repair man. I'm here to fix the air conditioner. Now, if you'd please excuse me. I've got work to do."
A 24 year old store cleark who received a gunshot wound (during the course of a robbery) which resulted in destruction of the right inferior convexity and orbital areas, attributed his hospitalization to a plot by the government to steal his inventions (Joseph 2007a). He claimed he was a famous inventor, had earned millions of dollars and had even been on TV. When it was pointed out that he had undergone surgery for removal of bone fragments and the bullet, he pointed to his head and replied, "that's how they are stealing my ideas."
Another patient, formerly a janitor, who suffered a large right frontal subdural hematoma (which required evacuation) soon began claiming to be the owner of the business where he formerly worked (Joseph 1988a). He also alternatively claimed to be a congressman and fabulously wealthy. When asked about his work as a janitor he reported that as a congressman he had been working under cover for the C.I.A. Interestingly, this patient, also stated he realized what he was saying was probably not true. "And yet I feel it and believe it though I know it's not right."
Frontal lobe confabulation seems to be due to disinhibition, difficulties monitoring responses, withholding answers, utilizing external or internal cues to make corrections, accessing appropriate memories, maintaining a coherent line of reasoning, or suppressing the flow of tangential and circumstantial ideas (Fischer et al. 2013; Joseph 2007a,1988a, 2011a; Johnson, O'Connor and Cantor 2007; Kapur and Coughlan 1980; Shapiro et al.1981; Stuss et al. 1978; Stuss and Benson 2007). That is, since the right frontal lobe can no longer regulate information processing and the flow of perceptual and ideational activity, information that is normally filtered out and suppressed is instead expressed. In consequence, the Language Axis of the left hemisphere becomes overwhelmed and flooded by irrelevant, bizarre associations, leading sometimes to the expression of false memories, which the patient (that is, Broca's area) repeats (Joseph, 1982, 2007ab, 1988ab).
As noted, in some respects injuries involving the orbital frontal lobes can result in symptoms similar to those with right frontal injuries, including the production of confabulatory ideation. However, in contrast to right (or bilateral) frontal injuries which may result in the production of fantastical spontaneous confabulations where contradictory facts are ignored or simply incorporated, confabulatory responses associated with orbital injuries tend to be more restricted, transitory, and in some cases must be provoked (Fischer et al. 2013).
DEPRESSION, APATHY, APHASIA, SCHIZOPHRENIA
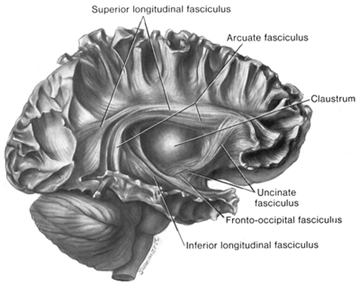
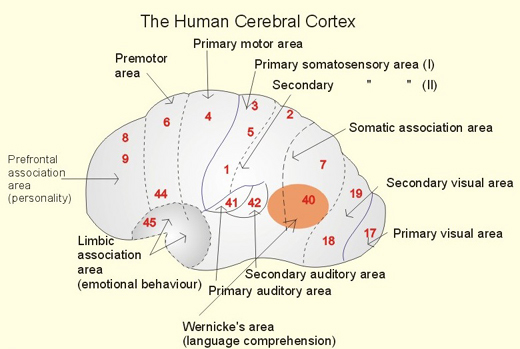
Broca's speech are is located in the general vicinity of the posterior- inferior region of the left frontal area (i.e. third frontal convolution), and includes portions of areas 45, 6, 4, and all of area 44. This region is multimodally responsive (Passingham, 2007) and receives projections from the auditory, visual, somesthetic areas (Geschwind, 1965; Jones & Powell, 1970), as well as massive input from the inferior parietal lobule and Wernicke's area via a rope of nerve fibers referred to as the arcuate fasciculus (which also links these areas to the amygdala). In addition, Broca's area receives fibers from and projects to the anterior cingulate as well as the brainstem periaqueductal gray which subserves vocalization.
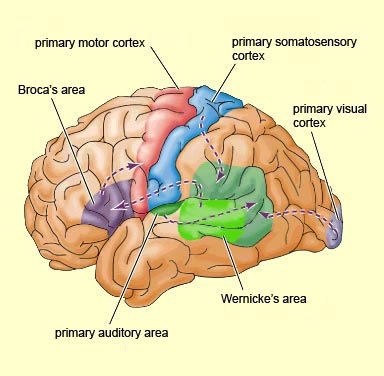
Broca's speech area is a final converging destination point through which thought and other impluses come to receive their final sequential (syntactical, grammatical) inprint so as to become organized and expressed as temporally ordered motoric articulations; i.e. speech (Foerster 1936; Fox 2013; Goodglass, 1993; Goodglass & Kaplan, 2000; Kimura, 1993; LeBlanc 1992; Petersen et al. 1988, 1990; Sarno, 1998). Verbal communication, the writing of words (via transmission to Exner's area), and the expression of thought in linguistic form is made possible via Broca's area which programs the adjacent oral-laryngeal musculature as represented within the adjacent primary motor areas (Foerster 1936; Fox 2013; Kimura, 1993; LeBlanc 1992; Petersen et al. 1988, 1990; Sarno, 1998); and which transmits to Exner's writing area so that words may be written.
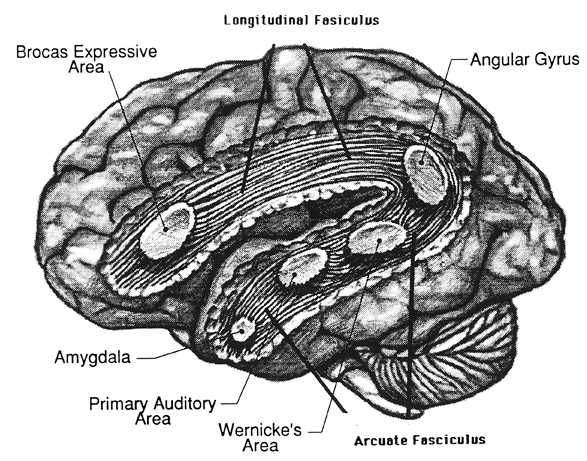
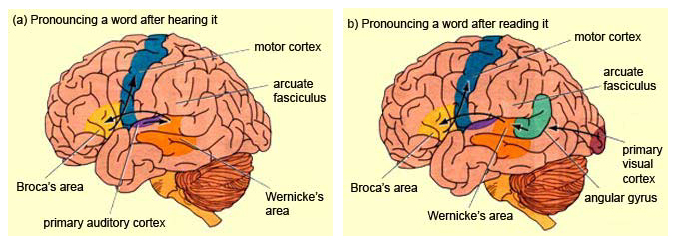
The importance of Broca's area and the left frontal lobe has also been demonstrated through functional imaging. For example, the left frontal lobe becomes activated during inner speech and subvocal articulation (Paulesu, et al., 1993; Demonet, et al., 2014). The left frontal lobe also becomes highly active when reading concrete and abstract words (Buchel et al., 1998; Peterson et al., 1988), and when engaged in semantic decision making tasks (Demb et al., 2013; Gabrielli et al., 2006). Moreover, activation increases as word length increases and in response to long and umfamiliar words (Price, 2007).
With injuries to Broca's area the individual loses the capacity to produce fluent speech (Goodglass, 1993; Goodglass & Kaplan, 2000; Sarno, 1998). Output becomes extremely labored, sparse, and difficult, and they may be unable to say even single words, such as "yes" or "no". Often, immediately following a large stroke patients are almost completely mute and suffer a paralysis of the upper right extremity as well as right facial weakness (since these areas are neuronally represented in the immediately adjacent area 4). Patients are also unable to write, read out loud or repeat simple words. Interestingly, it has been repeatedly noted that almost immediately following stroke some patients will announce "I can't talk", and then lapse into frustrated partial mutism.
With less severe forms of Broca's (also referred to as expressive, motor, nonfluent, verbal) aphasia, speech remains labored, agrammatical, fragmented, extremely limited to stereotyped phrases ("yes", "no", "shit", "fine") and contaminated with syntactic and paraphasic errors i.e. "orroble" for auto, "rutton" for button (Bastiaanse, 2013; Goodglass, 1993; Goodglass & Kaplan, 2011; Haarmann & Kolk 2014; Hofstede & Kolk 2014; Levine & Sweet, 1982, 2003; Sarno, 1998; Tramo et al. 1988). Writing remains severely effected, as are oral reading and repetition. Such patients also have mild difficulties with verbal perception and comprehension (Hebben, 2007; Maher et al. 2014; Tramo et al., 1988; Sarno, 1998; Tyler et al. 2013), including the ability to follow 3-step commands (Lura 1980). Commands to purse or smack the lips, lick, suck, or blow are often, but not always, poorly executed (DeRenzi et al. 1966; Kimura 1993); a condition referred to as bucal-facial apraxia. Non-speech oral movements are seldom significantly effected (Goodglass & Kaplan, 2011; Hecaen & Albert, 1978; Kimura 1993; Levine & Sweet, 2003).
With mild damage, patients may demonstrate severe confrontive naming and word finding difficulties (anomia), as well as possible right facial, hand, and arm weakness. Speech is often characterized by long pauses, misnaming, paraphasic disturbances and articulatory abnormalities (Goodglass, 1993; Goodglass & Kaplan, 2011; Sarno, 1998). Stammering and the omission of words may also be apparent. Similarly, electrical stimulation of this region results in speech arrest (Ojemann & Whitaker, 1978; Lesser et al., 1984) and can alter the ability to write and/or perform various oral-facial movements. It is noteworthy that even with anterior lesions or surgical frontal lobectomy sparing Broca's area a considerable impoverishment of spontaneous speech can result (Luria, 1980; Milner, 1971; Novoa & Ardila, 1987).
Disturbances involving grammar and syntax, and reductions in vocabulary and word fluency in both speech and writing have been observed with frontal lesions sparing Broca's area (Benson, 1967; Crockett, Bilsker, Hurwitz, & Kozak, 2007; Goodglass & Berko, 1960; Milner, 1964; Novoa & Ardila, 1987; Petrie, 1952; Samuels & Benson, 1979; Stuss, et al., 1998; Tow, 1955). In word fluency tests, however, simple verbal generation (e.g. all words starting with L) is usually more severely impaired than semantic naming, e.g. all animals which live in the jungle--which is presumably a function of semantic processing being more dependent on posterior language areas (Stuss et al., 1998).
DEPRESSION, APHASIA, & APATHY
Depression, "psycho-motor" retardation, apathy, irritability, and blunted mental functioning are associated with neocortical injuries of the left lateral and medial frontal lobe. When Broca's area has been injured, patients not only have difficulty with expressive speech (Bastiaanse 2013; Goodglass and Kaplan 2011; Haarmann and Kolk 2014; Hofstede and Kolk 2014; Sarno, 1998) but they typically become exceedingly frustrated, irritable, and depressed (Gainnoti 1972; Robinson and Benson 1981; Robinson and Szetela 1981; Robinson and Downhill 2006).
In large part, depression is common with Broca's aphasia as patients are painfully aware of their deficit (Gainotti 1972; Joseph, 1988a). Indeed those with the smallest frontal convexity lesions often become the most depressed (Robinson and Benson 1981). Depression in these cases appears to be a normal reaction and as such is mediated by undamaged tissue; i.e., the right hemisphere which is dominant for emotional expression and perception (e.g. Borod 1992; Cancelliere and Kertesz 1990; Freeman and Traugott 1993; Heilman and Bowers 2006; Joseph 1988a; Van Strien and Morpurgo 1992). That is, the right hemisphere being emotionally astute, reacts appropriately to the patient's condition and becomes depressed. If fact, with the exception of at least one study (Mayberg et al., 2011) almost all other studies demonstrate increased right frontal activity in response to negative moods (Rauch et al., 2006; Shin et al., 2007, 2011; Teasdale et al., 2011) and decreased left frontal activity with depression (Bench et al., 2013). In fact, repetitive transcranial magnetic stimulation of the right frontal lobe reduces depressive symptoms (Klein et al., 2011), whereas left frontal activity increase with the alleviation of depression as demonstrated through functional imaging studies (Bench et al., 2013).
Not only are left frontal injuries associated with tearfulness, irritability, and depression (where it is the right which may actually feel sad), but psychiatric patients classified as depressed, and normal individuals made to feel severely depressed, demonstrate insufficient left frontal activation and arousal (d'Elia and Perris 1973, Perris 1974; Tucker et al. 1981). For example, reduced bioelectric arousal over the left frontal region has been reported following depressive mood induction (Tucker et al. 1981). Similarly depressed mothers and depressed children show reduced left relative to right frontal activation (reviewed in Dawson 2014). With recovery from depression left hemisphere arousal returns to normal levels.

Likewise, Patients who are severely depressed have been shown to demonstrate insufficient activation and a significant lower integrated amplitude of the EEG evoked response over the left vs right frontal lobe (d'Elia and Perris 1973, Perris 1974). Based on EEG and clinical observation, d'Elia and Perris have argued that the involvement of the left hemisphere is proportional to the degree of depression. Moreover, with recovery the amplitude of the evoked response increases to normal left hemisphere levels. Functional imaging of depressed states indicates reduced activity in the left frontal lobe and anterior cingulate (Bench, et al,., 1992) and when these individuals ceased to be depressed, activity levels increases (Bench et al., 2013).
Left frontal lobe depression is therefore seen in those who are aphasic, and those whose depression has been long standing or even recently provoked. The more severe the depression, the greater is the reduction in left frontal functioning (whereas with mild transient sadness there might be a reduction in right frontal activity). Hence, with massive left frontal dysfunction, including even when Broca's area is spared, patient may become exceeding depressed, apathetic , hypoactive and indifferent (Robinson et al. 1984; Robinson and Szetela 1981; Sinyour, et al. 2007). However, with severe injuries, instead of worried or emotionally depressed, the patient instead is indifferent, uncaring, apathetic, and emotionally blunted (Blumer and Benson 1975; Freeman and Watts 1942, 1943; Girgis 1971; Hecaen 1964; Luria 1980; Passingham 1993; Stuss and Benson 2007; Strom-Olsen 1946). One patient who "prior to his accident requiring amputation of the left frontal pole, had been garrulous, enjoyed people, had many friends, was active in community affairs" and had "true charisma... became quiet and remote, spent most of his time sitting alone smoking, and was frequently incontinent of urine, and occasionally of stool. He remained unconcerned and was frequently found soaking wet, calmly sitting and smoking. When asked, he would deny illness" (Blumer and Benson 1975, p. 196).
Similarly, inertia and apathy usually imediately follow surgical destruction or injury of the frontal lobes, the left frontal lobes in particular (Hillbom 1951; Lishman 1968). "The previously busy housewife who has always been a dirt-chaser, and who has kept her fingers perpetually busy with darning, crocheting, knitting, and so on, sits with her hands in her lap watching the 'snails whiz by'. Like a child she must be told to wipe the dishes, to dust the sideboard, to sweep the porch" and even then the patient completes only half the task as there is no longer any interest or initiative (Freeman and Watts 1943, p. 803). In some cases the apathy is so profound that "whoever has charge of the patient will have to pull him out of bed, otherwise he may stay there all day. It is especially necessary since he won't get up voluntarily even to go to the toilet" (p. 802).
Depressive-like features, however, also seem to result with left anterior damage sparing Broca's area such as when the frontal pole (of either hemisphere is compromised (Robinson et al. 1984; Robinson & Szetela, 1981; Sinyour, et al. 2007). As described by Kennard (1939) monkeys with bilateral frontal lobe damage would sit with their head sunk between their shoulders, neither blinking or turning their heads in response to noise, threats or the presence of intruders; but would stare absently straight ahead with no facial expression. In a similar study, following massive frontal destruction a monkey who was formerly quite active and the dominant leader of his group became inactive, indifferently watched others, failed to respond emotionally, and seemed to have lost all interest and ability to engage in complex social behavior (Batuyev, 1969).
In his summary of two large scale frontal tumor studies Hecaen (1964) noted the majority seemed confused, disorganized, apathetic, hypoactive, and suffering from inertia and feelings of indifference.However, puerility was also common among these patients and many demonstrated decreased judgment with either total or partial unawareness of the environment.
In some, this initial state of inertia disappears, whereas in others it becomes a lasting or even progressively severe disturbance. "The previously busy housewife who has always been a dirt-chaser, and who has kept her fingers perpetually busy with darning, crocheting, knitting, and so on, sits with her hands in her lap watching the 'snails whiz by'. Like a child she must be told to wipe the dishes, to dust the sideboard, to sweep the porch" and even then the patient completes only half the task as there is no longer any interest or initiative (Freeman & Watts, 1943, p. 803).
In part, depression coupled with apathy secondary to frontal injuries is probably related to damage to the interconnections with the medial region, an area which when damaged induces hypokinetic and apathetic states (see below). However, these latter patients are not depressed, but rather severly apathetic, indifferent, hypoactive, and poorly motivated. When questioned, rather than worried or truly concerned about their condition the overall picture is that of confusion, disinterest, and blunted emotionality (Freeman & Watts, 1942: Hacaen, 1964) i.e. there is a lack of worrisome thoughts or depressive ideation.
Hence, in part, apathetic and depressive features may result from left frontal convexity and frontal pole damage due to a severance of fibers which link emotional impulses (such as those being transmitted via the orbital and medial region) with external sources of input or cognitive activity which are transmitted to the convexity (i.e. disconnection), impulses which are transmitted from the medial frontal lobes to the orbital, superior, and anterior frontal lobes. Through these interconnections, emotional impulses arising in the limbic system, can be transmitted through the medial frontal lobes to the lateral frontal lobes, where they then become ideas. Or conversely, neocortical cognitions may be transmitted from the lateral neocortical surface to the medial areas where they are integrated to again become emotional ideas.
It was the recognition that the frontal lobes acted as a bridge between emotion and idea which led to the wide scale use of frontal lobotomy; i.e. surgical destruction of inter-linking fibers --a technique, which when used in the 1940s and 1950s, often involved little more than blindly swishing a "surgical ice pick" inside somebody's brain!
Moreover, convexity lesions, like medial damage, may result in a disconnection not only between cognitive-perceptual and emotional activity, but would prevent limbic system output from reaching the motor areas such that emotional-motivational impulses are unable to become integrated with motor activities. The patient is thus motorically hypo-emotionally aroused (i.e. depressed), and appears to be demonstrating psychomotor retardation. Just as left frontal convexity motor damage can result in left sided apraxia (due to right hemisphere disconnection from left parietal temporal-sequential output), the reverse can also occur. That is, with left frontal damage, linguistic impulses not only fail to become expressed, but emotional output from the right hemisphere and limbic system fail to become integrated with linguistic-ideation (i.e. thought). Ideas no longer come to be assigned emotional significance. In the extreme the motivational impetus to even engage in thought production is cut-off.
As pertaining to laterality, left frontal (vs. right) lesions are associated with reductions in intellectual and conceptual capability which often leads to confusion and a reduced ability to appreciate and appropriately respond to the external or internal environment. In these instances, one possible consequence is apathy, indifference, hyporesponsiveness, and depressive-like symptoms.
On the otherhand, it is possible that among psychiatric patients and otherwise normal, albeit, depressed individuals, that the left frontal region appears relatively inactive because the right frontal area is preoccupied with being depressed; the right frontal region is excessively aroused (Teasdale, et al., 2011). That is, excessive right frontal arousal leads to massive left frontal inhibition; i.e., the bilateral arousal system of the right hemisphere inhibiting the left.

PSYCHOSIS & BLUNTED (NEGATIVE) SCHIZOPHRENIA
Patients with left frontal injuries or dysfunction, and who become unresponsive, apathetic, untidy, and whose speech is abnormal, are sometimes characterized as suffering from an emotionally blunted form of schizophrenia. In fact, left lateral as well as bilateral convexity abnormalities are often associated with apathetic, blunted, and "negative" forms of schizophrenia (Buchsbaum 1990; Carpenter et al. 1993; Casanova et al. 1992; Weinberger 1987; Wolkin et al. 1992). Similarly, patients classified as schizophrenic have also been reported to demonstrate abnormal left( or bilateral) frontal lobe EEG's indicative of hypoarousal (Akbarian et al. 1993; Ariel et al. 2003; Ingvar and Franzen 1974; Kolb and Whishaw 2003; Levin 1984). Lateral frontal gray matter and brain volume reductions and decreased activity have been repeatedly noted (Andreasen et al. 1990; Buchanan et al. 1998; Curtis et al. 1998), including decreased blood flow (Weinberger et al. 2007) hypoactivity (Wolkin et al. 1992), reduced metabolism (Buchsbaum et al. 1992), as well as left sided abnormalities affecting the striatum (Breier et al. 1992; Buchanan et al. 1993; Swayze et al. 1992) which is buried within the depths of the frontal lobe.
This is not to imply that all subtypes of "schizophrenia" are secondary to frontal lobe pathology. The left temporal lobe have also been implicated as measured by positron-emission tomography (e.g. McGuire et al. 1998), P300 evoked potential amplitude (Bruder et al., 2011; Salisbury et al. 1998) and MRI (Jacobsen et al. 1998; Shidhabuddin et al. 1998; Kwon et al., 2011). However, as the temporal lobes are implicated in these subtypes of schizophrenia, the pattern of symptoms differs from those with left frontal dysfunction.
For example, depending on if the right vs left and superior vs the inferior temporal lobe are more greatly impacted, patients are more emotional, more verbal, more active, and more likely to suffer visual and auditory hallucinations, coupled with disturbances of comprehension. However, whereas as a superior temporal lobe (Wernicke's area) abnormality can affect the left frontal lobe (Broca's area) thus producing fluent-aphasic speech and thus a complete formal thought disorder, left inferior temporal lobe and amygdala dysfunction can disrupt the orbital and medial frontal lobe including the caudate and putamen. Indeed, the striatum evolved from and is densely interconnected with the amygdala, and if injured mental functioning becomes invariably abnormal.
Hence, it appears that only certain subpopulations of schizophrenics actually suffer from frontal lobe dysfunction, such as those with catatonia, posturing, mannerisms, and emotional blunting coupled with reduced speech output and apathy. However, not all "frontal" schizophrenics are blunted, but may display unusual mannerisms and/or a silly, puerile childishness that long ago had been referred to as "Hebephrenia" Indeed, Hillbom (1951), described a number individuals with head trauma and missle wounds to the frontal lobes who developed schizophrenic-like symptoms, including catatonia and hebrephrenia. However, Hillbom (1951) also found that left frontal patients are more likely to develop these symptoms.
THE STRIATUM: EMOTION, PLEASURE, BODY LANGUAGE, AND SCHIZOPHRENIA
The corpus striatum (caudate and putamen) and limbic striatum (olfactory tubercle, nucleus accumbens, stria innominata) are buried deep within the frontal lobe, with the head of the caudate arising from near the orbital floor and then ascending deep alongside the inner medial and lateral walls (Mink, 2007)
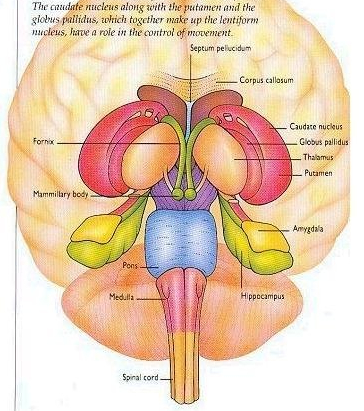
The striatal nuclei are thus intimately associated with the orbital, medial and lateral frontal lobes, as well as the amygdala with which they merge in the temporal lobe. It is through these frontal-striatal-amygdala pathways that emotional states come to translated into complex motor acts and expressed through body language (see Figure 5). That is, the striatum mediates facial expression and gesture, and produces ballistic movements, including hitting, kicking, running and biting (Mogenson and Yang 1991; MacLean 1990; Rapoport 1991).
Since humans possess basically the same striatum and limbic system, when happy, sad, angry, and so on, the facial and body musculature assumes the same readily identifiable emotional postures and expression regardless of culture or racial orgins (Ekman 1993; Eible-Ebesfedlt 1990; Joseph, 1993). The striatum is capable of considerable flexibility in motor-emotional expression, and is exceedingly responsive to the internal motivational and emotional state. Through its extensive interconnections with the limbic system and the mesolimbic dopamine (DA) system the striatum is able to perceive emotional stimuli and can "experience" pleasure. Striatal neurons selectively respond to motivationally significant stimuli that are rewarding or punishing (Rolls and Williams 1987; Schneider and Lidsky 1981) and can generate feelings of pleasure in reaction to amphetamine, cocaine and opiates (Ellison 2014; Hakan et al. 2014; Koob et al. 1991). Destruction of striatal neurons disrupts the capacity to experience pleasure such as mediated through opiates and cocaine (Koob et al. 1991).
Similarly, lesions to the corpus striatum and lenticular nucleus (putamen and globus pallidus) can attenuate one's capacity to experience pleasure or motorically express their emotions via the musculature; e.g. the face and body becomes frozen; a condition similar to Parkinson's disease. In fact, surgical destruction of or chemical lesions to the anterior corpus striatum and surrounding tissue can produce catatonic or "frozen" states, including prolonged posturing where the subjects will remain unmoving for hours, days, and even weeks at a time (Denny-Brown 1962; Spiegel and Szekely 1961).
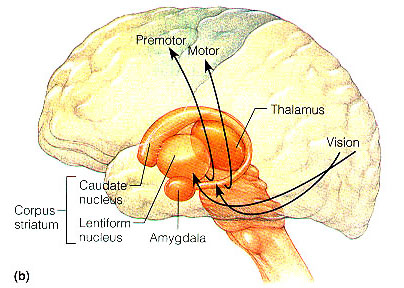
Similar paralytic, petrified disturbances can be induced with dopamine depletion. Indeed, a major function of striatal DA is inhibition (Ellison 2014; Le Moal and Simon 1991; Mercuri et al. 1985), and DA depletion can trigger striatal as well as limbic hyperactivation (chapter 30). If hyperactivated, associated muscular activity as well as the neural pathways linking the striatum with the frontal lobe and amygdala, can be disrupted and overwhelmed, thereby giving rise to emotional and behavior paralysis. The patient may become catatonic (see below) or in the less extreme may demonstrate signs of Parkinson's disease.
Likewise, excessive DA, and reduced striatal activity can disrupt all aspects of motor, cognitive and emotional functioning, with some patients developing an acute schizophrenic psychosis (Castellanos et al. 2014; Chakos et al. 2014; Ring et al. 2014; Shihabuddin et al. 1998; Snyder 1972). In addition to reduced activity and metabolism (Buchsbaum, et al. 1992; Resnick et al. 1988; Wiesel et al. 1987), as based on MRI, limbic striatal structures are reduced in size (Shihabuddin et al. 1998) whereas the corpus striatum has been found to be increased in size in schizophrenics vs normal controls (Heckers et al. 1991; Shihabuddin et al. 1998) especially in the left hemisphere (Breier et al. 1992; Buchanan et al. 1993; Swayze et al. 1992). Moreover, as the lateral frontal neocortex is also innervated by striatal as well as DA fibers, reduced DA levels can also negatively impact the lateral frontal lobes as well as the striatum (Beatty et al. 1993).
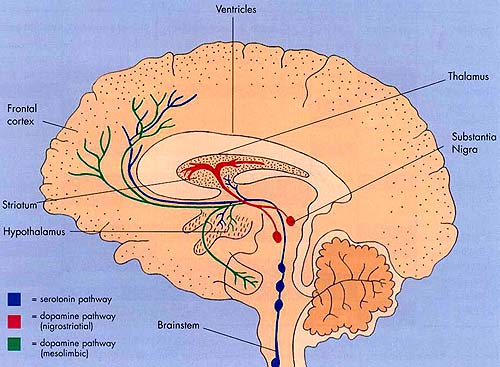
Striatal abnormalities are linked to frontal abnormalities and vice versa. In consequence, if the frontal lobes become abnormal secondary to striatal/DA disturbances, frontal symptoms will be evoked. Hence, striatal dysfunction is also associated with depression, apathy, obsessive compulsive disorders, "schizophrenia," as well as mania (Aylward et al. 2014; Baxter et al. 1992; Caplan, et al. 1990; Castellanos et al. 2014; Chakos et al. 2014; Deicken et al. 2013; Ellison 2014; Rauch et al. 2014; Richfield et al. 1987; Robinson and Downhill 2013), depending on the nature, degree, extent, and laterality of destruction.
For example, injuries or abnormalities restricted to the right caudate are more likely to result in a manic-like psychosis (Castellanos et al. 2014; Robinson and Downhill 2013) --similar to right frontal lobe injuries. By contrast, left or bilateral caudate injuries are associated with apathetic states, decreased spontaneous activity, slowed and delayed dysarthric and emotionally flat speech, with some patients responding to questions only after a 20-30 second delay (Caplan et al. 1990; Richfield et al. 1987).
For example, Richfield et al. (1987, p. 768) described a 25 year old female honor student, soon to be married, who suddenly disappeared after complaining of headaches and nausea. When found three days later she "had undergone a dramatic personality change manifested by alterations in affect, motivation, cognition, and self-care which included vulgarity, impulsiveness, violent outbursts, enuresis, indifference, hypersexuality, shoplifting and exposing herself. She was inattentive and uninterested in her surroundings. She would frequently lie down to sleep. Her affect was flat." CT-scan indicated bilateral damage to the head of the caudate nuclei as well as surrounding structures.
Since the head of the striatum descends ventrally between the lateral and medial walls, injuries to this part of the brain, including DA abnormalities, also disrupts these adjoining tissues. If the right or left lateral walls are injured, patients are likely to appear manic or apathetic, depressed, irritable, and schizophrenic. However, if the medial walls of the frontal lobes are injured, cognitive, motivational, and motor functioning will become profoundly abnormal, and the patient may become completely catatonic.
MOVEMENT AND THE MOTOR AREAS OF THE FRONTAL LOBES
FUNCTIONAL NEUROANATOMY OF THE FRONTAL MOTOR AREAS
Movement and motor functioning are dependent on the functional integrity of the basal ganglia, brainstem, cerebellum, cranial nerve nuclei, the motor thalamus, spinal cord, as well as the primary, secondary and supplementary motor areas of the frontal lobes (Passingham, 2007; Schmahmann, 2007). Indeed, these areas are all interlinked and function as an integrated system in the production of movement (Mink, 2007; Mink & Thach, 1991; Parent & Hazrati 2013; Passingham, 2007).
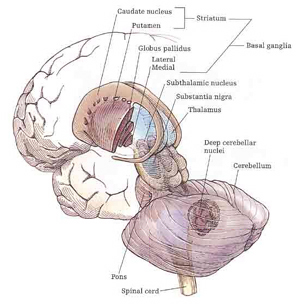
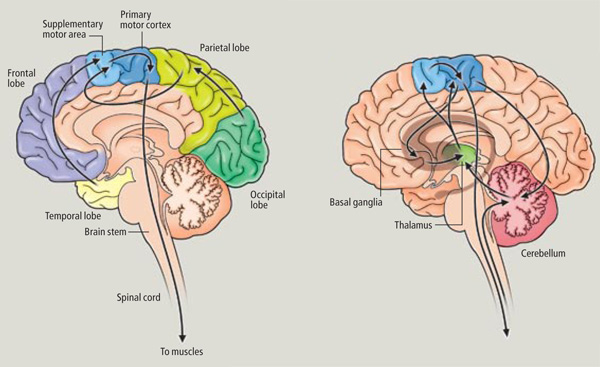
However, as to fine motor movements including those involved in the articulation of speech, these are almost completely dependent upon the functional integrity of the primary motor areas located along the percental gyrus (area 4), and within which are represented the muscles controlling the hands, fingers, and oral laryngeal musculature.
And yet, as noted above, the primary motor areas are in turn dependent upon motoric impulses which are organized in premotor and the supplementary motor cortex--the latter of which is located along the medial wall of the hemispheres. Considered from a very broad and simplistic perspective, it could be said that primary area is programmed and under the control of the secondary and supplementary motor areas as well as the "prefrontal" and other areas of the cerebrum, although neurons in the primary area also become active prior to and during movement (Passingham, 2007).
For example, Exner's writing area is in part, within areas 6 and becomes active prior to (as well as during) hand movements and appear to program hand movements, whereas the frontal eye fields (within areas 6,8,9) becomes active prior to (as well as during) eye movements and appears to program eye movements. As noted above, the primary area representing the oral-laryngeal musculature, is programmed by Broca's expressive speech area areas 45, 46 (Foerster 1936; Fox 2013; LeBlanc 1992; Petersen et al. 1988, 1989). Broca's area also becomes active prior to vocalization and during subvocalization as indicated by functional imaging.
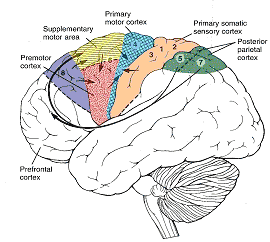
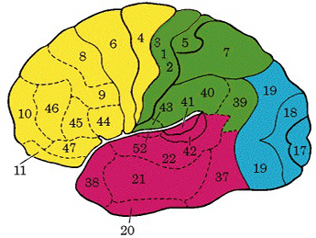
Hence, like the sensory areas where information is generally received in the primary zones before transmission to the association areas as well as in the association areas independently of the primary areas (Zeki, 2007) motor impulses are processed and acted on in parallel (Passinghma, 2007), though in general, they generally begin their organizational journey in the supplementary motor areas (Alexander & Crutcher, 1990; Crutcher & Alexander, 1990) and/or the cingulate and other forebrain structures (Passingham, 2007; Stephan, et al., 2011), which are then transmitted to the primary regions where they are acted upon.
Again, however, there are areas within the primary region which also become active prior to movement, though these same areas become increasing active during movement (Passingham, 1993).
Because, in general, the impulse to move originates outside the primary motor cortex, direct electrical stimulation of this region does not give rise to complex, coordinated, or purposeful movements (Penfield & Boldrey, 1937; Penfield & Jasper, 1954; Penfield & Rasmussen, 1950; Rothwell et al. 1987). Rather stimulation will only induce, for example, twitching of the lips, flexion or extension of a single finger joint, protrusion of the tongue or elevation of the palate. It is noteworthy that patients never claim to have willed these movements, which again suggests the will-to-move is initiated elsewhere.
However, if electrical stimulation is applied when the patient is attempting to move, the result is paralysis (Penfield & Jasper, 1954; Penfield & Rasmussen, 1950). Presumably the reception of impulses-to-move (which are initiated and organized elsewhere) are blocked by primary motor electrical stimulation.
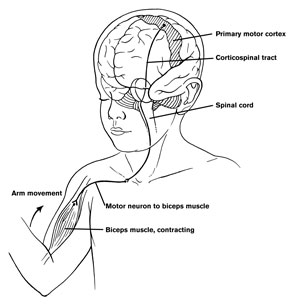
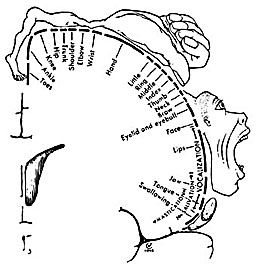
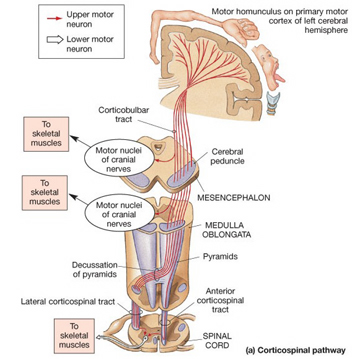
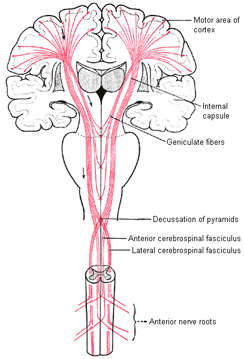
Motor functioning and movement, however, is also dependent on the parietal lobes, thalamus, basal ganglia, brainstem, cerebellum, and spinal cord, structures which are directly or indirectly interconnected (Kaas, 1993; Mink, 2007; Passingham, 2007; Schmahmann, 2007).
For example, smooth, purposeful, coordinated movement requires sensory feedback which is provided by the primary and association/secondary somatosensory areas located in the parietal lobe and which contain neurons which guide the hand and arm in visual space (see chapter 20). These cortices are intimately linked with the primary motor and the motor association/secondary and supplementary motor areas as well as the basal ganglia, brainstem, and cerebellum--regions which become highly active during and often prior to movement initiation (Passingham, 2007). For example, when human subjects learn a sequence of finger movements, functional imaging reveals increased activity in the primary and premotor cortex, and the primary and association somesthetic cortex, as well as in the cerebellum, striatum, ventral thalamus and cerebellum (Passingham, 2007). In fact, most of the "motor" area also contain independent motor maps of the body, which is why, if a subject moves an arm, leg, or shoulder, each of these areas becomes active almost simultaneously--at least as demonstrated with functional imaging.
On the other hand, as based on functional imaging, it has also been shown that there is increased activation of the primary motor area when subjects actually make a movement, as compared to merely preparing to make the movement (Passingham, 2007).
Hence, although some movements are programmed in parallel by a number of motor areas simultaneously, others are programmed in temporal sequences such that the SMA programs the premotor area which acts on the primary area which becomes maximally active when movements are actually performed. IN this regard it is noteworthy that whereas stimulation of the SMA can prevent movement, stimulation of the premotor area can give rise to the illusion that a movement is about to be made, that is, of an impending movement (Penfield, 1938), whereas activation of the primary area can trigger twitching, and minor motor movements--which again suggests a hierarchical organization.
According to Carpenter et al., (2011, p. 1755), although "individual cells prove largely independent information about the items in the sequence... during different epochs of presentation of the stimuli, different patterns of distributed activity in even small ensembles of motor cortical cells are sufficiently distinct and robust to provide a basis for encoding the sequence," which in turn allows for memory scanning and retrieval.
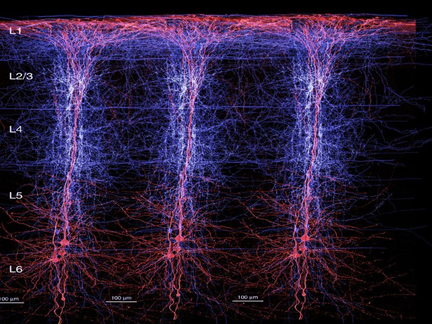
In addition, "motor" neurons in the inferior-ventral portion of the (monkey) premotor cortex can code for the spatial location of nearby sounds, particularly those originating near the head (Graziano et al., 2011). Indeed, these neurons appear to be "multi-modal" and can also respond to visual and tactile stimuli that impinge near or upon the face (Graziano et al., 2011).
Only approximately 31% of the corticospinal tract arises from the pyramidal cells of agranular primary motor area 4, with another 30% arising from areas 6, 8, and the SMA, and another 30% from the primary and secondary/association somesthetic areas in the parietal lobe, and the remainder arising from the deep layers of the temporal, occipital, and orbital frontal lobes, as well as from pyramidal cells scattered throughout the amygdala, cingulate, and striatum. Hence, widespread areas of the neocortex control or influences movement by acting directly on the brainstem and spinal cord via the corticospinal tracts.
Presumably, those corticospinal axons arising from the parietal lobe project to the dorsal horns of the spinal cord so as to regulate sensory inflow rather than directly controlling movements. Hence, as two thirds of the corticospinal tract arise in the frontal motor area as these project to motor neurons, it is the frontal motor areas which dominate and exert controlling influences over most aspects of purposeful motor behavior. Because the motor cortex is specialized for controlling movement, the functional organization of the motor cortex differs from that of other brain areas.
For example, the neocortex of the motor areas are relatively thick and "agranular" in structure. This is because internal "granular" layer IV is obscured by the increased size of layers III and V which are packed with large numbers of exceedingly large pyramidal cells including the giant cells of Betz. By contast, in somesthetic areas 3,1,2, layers III and V are much smaller, whereas layer IV is much thicker.
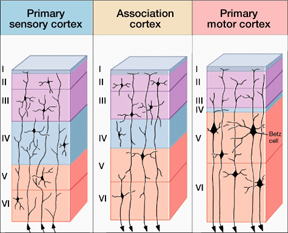
Within the frontal motor areas, it is layers III and V which gives rise to the corticospinal tract, and which consists of large and medium size pyramidal cells. Layer V also gives rise to the corticbulbar, corticopontine, and corticorubral fibers.
Approximately 90% of corticospinal axons range from 1 to 4 um in diameter. The largest fibers are axons from the giant Betz cells which are located in layer V of the primary motor area. About 60% of the axons of the corticospinal tract are myelinated.
Specifically, the axons of the corticospinal (and related) tracts arise from large pyramidal neurons located in layers III and V of the neocortex and descend via the internal capsule to the midbrain, some of which make contact and terminate in the red nucleus, the periaqueductal gray, and a variety of other brainstem motor nuclei. Collectively these latter fibers are actually referred to as the corticobulbar tract and many of its axons arise in the orbital and medial frontal lobe as well as the inferior lateral convexity.
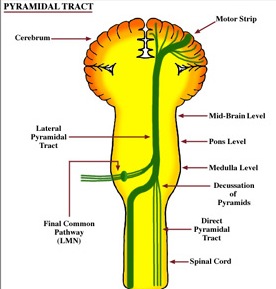
By contrast, corticospinal axons descend through the pons where they separate into tiny nerve bundles before regrouping within the medulla to form the medullary pyramid. However, at the spinal-medulla border, about three fourths of these axons cross over at the midline of the medulla to form the pyramidal decussation, with the crossed fibers forming the lateral corticospinal tracts and the remaining uncrossed axons forming the ventral corticospinal tract.
Hence, the corticospinal tract aconsists of two divisions, the lateral and ventral.The lateral tract projects to the lateral motor nuclei of the ventral horn and to intermediate zone interneurons, whereas the ventral tract projects bilaterally to the medial cell column which is concerned with the axial muscles. Most of the ventral (uncrossed) axons originate in Broadmans areas 4, 6 and 8, whereas the lateral tract also originates in frontal motor areas 4 and 6, and parietal somesthetic areas 3,1,2,5,7, though yet others have their source throughout the neocortex as well as in the limbic system.
Thus, these two divisions consist of a number of separate fiber bundles which originate not just within the pyramidal cells of the somatomotor areas, but within the striatum, cingulate and amygdala. These fiber bundles, therefore consist of a commingling of axons which originate throughout the forebrain, and which arise in discrete neocortical locations. Again, widspread areas of the brain contribute to purposeful movement. Moreover, although these fiber bundles descend together through the internal capsule, they innervate different brainstem targets, and are thus referred to as the corticobulbar, corticopontine, corticorubral and corticospinal tracts.
The vast majority of descending motor fibers, however, penetrate the brainstem and establish direct synaptic interconnections with spinal motor nuclei. These latter fibers are referred to as the corticospinal tracts.
Specifically, corticospinal axons descend through the pons where they separate into tiny nerve bundles before regrouping within the medulla to form the medullary pyramid. At the spinal-medulla border, about three fourths of these axons cross over at the midline to form the pyramidal decussation, with the crossed fibers forming the lateral corticospinal tracts, and the remaining uncrossed axons forming the ventral corticospinal tract. That is, about 90% cross over, such that the left half of the brain controls the right half of the body (and vice versa).
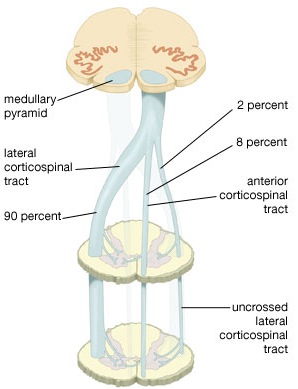
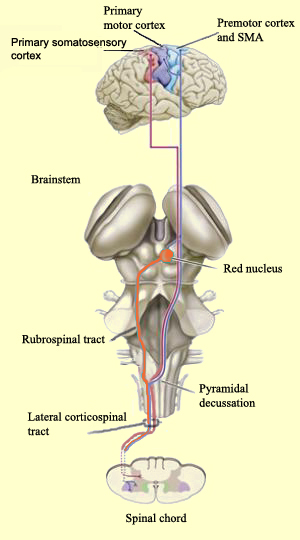
By these long descending pyramidal axons, neocortical motor control is gained via neocortical pyramidal axons which innervate the motor nuclei of the brainstem and spinal cord at all levels.
Hence, due to early left hemispheric motor development, the left corticospinal tract grows more quickly and descends into the brainstem, and then crosses at the pyramidal decussation, and then descends into the spinal cord in advance of those fibers from the right (Kertesz & Geschwind 1971; Yakovlev & Rakic 1966). This provides the left hemisphere and the right hand with a competitive advantage in establishing both motor control and thus right hand dominance.
Because the corticospinal tract does not reach advanced levels of myelination and does not become functionally mature until the 8th-12th postnatal month (Yakovlev & Lecours, 1968) it is only as the infant approaches their first birthday that handedness become obvious or apparent.
It is also around the 8th to 12th month that the motor and somatosensory areas become increasingly mature (Bell & Fox, 2014; Brody et al., 1987; Chugani, 2014; Hudspeth & Pribram, 1992; Huttenlocher, 1990; Schore, 2014) and the secondary motor and sensory receiving areas display increased myelination (Flechsig, 1901).
As based on PET scans, the frontal lobes also display increased metabolic activity from 8 to 12 months of age (Chugani, 2014), and around the end of the first year the neocortical EEG begins to increasingly resemble a more adult pattern (Kellaway, 1979; Ohtahara, 1981). In addition, around the 8th to 12th month, the corticospinal and pyramidal axons begin to approximate adult level of cellular development, myelination, and organization, and then become increasingly well myelinated over the course of the second year (Debakan, 1970; Langworthy, 1937; Yakovlev & Lecours, 1967)--a process which continues into late childhood (Paus et al., 2011).
Hence, around the 8-12th month the neocortex and frontal motor areas begin to increasingly exert hierarchical control over the limbic forebrain, brainstem, and musculature, and this process continues to develop over the course of the ensuing decade and requires considerable coordinated interaction between the parietal and motor areas of the frontal lobe as well as the striatum, limbic system, and cerebellum.
The SMA is tightly interlinked with the striatum, anterior cingulate, amygdala, as well as the motor thalamus, and contributes axons to the corticospinal tract and makes synaptic contact with the brainstem, cerebellum, and spinal cord (Brodal, 1981; Carpenter, 1991; Schmahmann, 2007). Moreover, the SMA receives input from and transmits to the premotor and primary frontal motor areas as well as from and to the parietal lobe. Hence, this structure is situated at the cross-roads where divergent impulses and sensory impression are received and apparently acts to integrate these signals. Presumably because of its integrating role, the SMA also becomes highly active during the modification, learning and establishment of new movement programs (Brinkman & Porter, 1979; Roland et al.1980; Tanji et al. 1980), and may well act to make decisions regarding possible movements based on these signals well before a movement is initiated.
In fact, single cell recordings (Brinkman & Porter, 1979; Tanji & Kurata, 1982) and studies of blood flow studies (Orgogozo & Larsen, 1979; Shibasaki et al. 1993) and movement related evoked potentials (Ikeda et al. 1992) indicate increased activity in this area when simply imagining as well as when performing complex movements of the fingers and hands.
In general, however, and as based on single cell recording (vs functional imaging) activity begins in the SMA well before movements are initiated and prior to activation within the premotor and primary motor areas (Alexander & Crutcher, 1990; Crutcher & Alexander, 1990; Mink, 2007; Mink & Thach, 1991). For example, when anticipating or preparing to make a movement, but prior to the actual movement, neuronal activity will first begin and then dramatically increase in the SMA, followed by activity in the premotor and then the primary motor area, and then the caudate and last of all the putamen and globus pallidus (Alexander & Crutcher, 1990; Mink, 2007; Mink & Thach, 1991).
Presumably the putamen, in conjunction with the caudate, transmits this information to the globus pallidus which in turn projects to the motor thalamus, brainstem reticular formation, as well as to the motor neocortex, thus creating a very elaborate feedback loop (Mink, 2007; Mink & Thach, 1991; Parent & Hazrati 2013) which is also influenced by the anterior cingulate and amygdala.
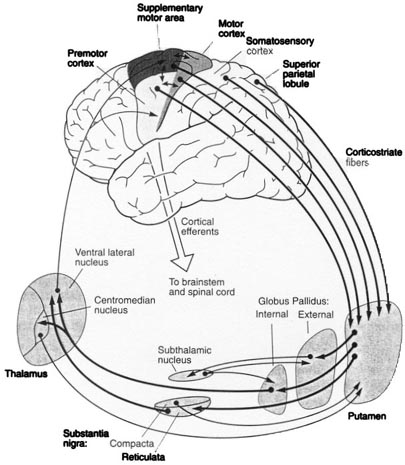
The SMA, however, is not concerned with fine motor functioning, as this is the province of the primary motor areas. Rather, it is concerned with coordinating gross and bilateral movements of the hands and extremities, including grasping functions (Andres et al., 2011; Passingham, 2007; Stephan et al., 2011). Electrical stimulation of the SMA has produced complex semipurposeful movements, vocalization (Penfield & Jasper, 1954), and postural synergies involving the trunk and extremities bilaterally (Van Buren & Fedio, 1976), but not fine motor movements.
Because of its role in guiding and programming gross body movements, if this structure is damaged, the body may become stiff and movements tend to be slow and incoordinated--a condition also seen with mild lesions (Penfield & Jasper, 1954). Patients demonstrate clumsiness, severe agraphia, impairments of bimanual coordination and difficulty performing rapid or alternating movements (Brinkman, 1981; Gasquione 1993; Goldberg & Bloom 1990; Goldberg et al. 1981; McNabb et al. 1988; Penfield & Jasper, 1954; Stephan et al., 2011; Travis, 1954; Truell et al. 2013; Watson et al. 2007). Patients may walk with short steps and suffer disturbances involving posture, balance, and gait. As detailed above, with extensive and massive injuries patients may become mute (McNabb et al., 1988; Watson et al., 2007) and so stiff and unmoving that they appear to demonstrate all the classic signs of catatonia including gegenhalten and waxy flexibility.
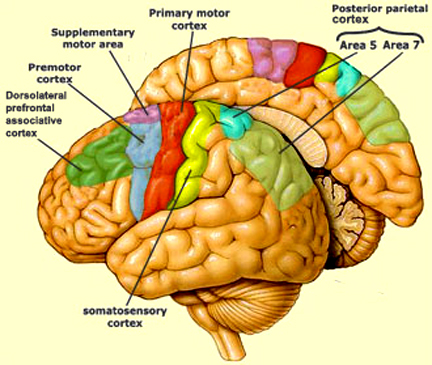
In addition to its interconnections with the SMA and primary motor areas (Jones et. al., 1978; Jones & Powell, 1970), the premotor area receives information directly from the primary and secondary somesthetic and visual (area 17, 18, 19) cortices (Jones & Powell, 1970; Pandya & Kuypers, 1969). It is precipally concerned with the guidance and refinement of movement via the assimilation of sensory information provided by the sensory areas (Godschalk et al. 1981; Porter 1990) and interacts with the basal ganglia, motor thalamus, and SMA so as to achieve these goals (Alexander & Crutcher, 1990; Crutcher & Alexander, 1990; Mink & Thach, 1991; Parent & Hazrati 2013).
As noted, the SMA becomes activated prior to the premotor area, which becomes activate prior to the primary motor area. Hence, whereas neurons in the primary motor region become active during movement, excitation in the premotor cortex precedes cellular activation of the primary region (Weinrich et al. 1984). Moreover, cells in the premotor cortex become activated before movements are even initiated, whereas electrical stimulation of this area induces the illusion of a impending movement (Penfield, 1938). These and other findings suggest that the premotor area may be modulating and exerting controlling and integrative influences on impulses which are to be transmitted to the primary region for expression.
Indeed, the premotor area appears to be highly involved in the programming of various gross and fine motor activities, and becomes highly active during the learning of new motor programs (Passingham, 2007; Porter 1990; Roland et al. 1981). Moreover, electrical stimulation elicits complex patterned movement sequences as well as stereotyped and gross motor responses such as head turning or torsion of the body (Fulton, 1934; Passingham, 1981, 1993).
Unlike the primary area, damage limited to the pre-motor cortex does not result in paralysis but disrupts fine motor functioning and dexterity, including simple activities such as finger tapping (Luria, 1980). With extensive damage fine motor skills are completely lost and phenomena such as the grasp reflex are elicited (Brodal, 1981); i.e. if the patient's hand is stimulated it will invulantarily clasp shut.
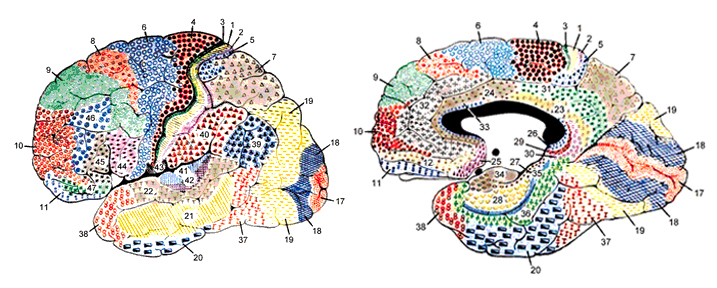
Functionally and structurally the primary motor cortex appears to extend well beyond the confines of the precentral gyrus (area 4) and includes portions of area 6 and the somatosensory (1, 2, 3, 5) regions (Brodal, 1981; Carpenter, 1991). These areas are richly interconnected (Jones et al. 1978; Jones & Powell, 1970; Kuypers & Catsamn-Berrevoets, 1984), the somatosensory projections providing information important in the sensory guidance of movement (Godschalk et al. 1981; Kaas, 1993; Lebedev et al. 2014).
As noted, axonal projections from the motor and somesthetic regions all give rise to the massive cortico-spinal (pyramidal) tract which innervates cranial nerve and sensory and spinal motor neurons (Brodal, 1981; Kuypers & Catsman-Berrevoets, 1984). For these reasons some authors have referred to the somesthetic and motor regions as the sensorimotor cortex.
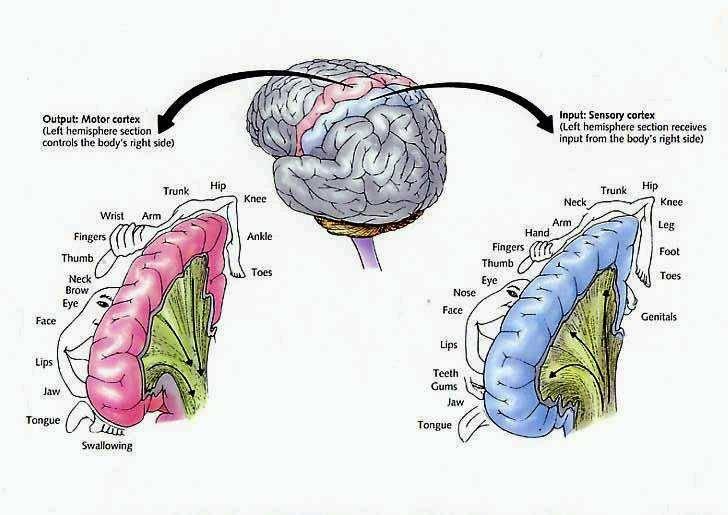
The primary motor area is concerned with the coordination and expression of gross and fine motor functioning including finger movements (Colebatch et al., 1991; Luria 1980; Rao et al. 2013; Sanes et al., 2013; Shibasaki et al. 1993; Woolsey, 1958), and serves as the final common pathway where impulses organized in other brain areas are transferred for expression, particularly those involving the fingers, thumb, lips, tongue, and hand. Indeed, the region of area 4 that represents the hand is especially prominent and in fact forms a bulge upon the surface (Yousry et al., 2013).
Electrical stimulation of discrete points within the motor cortex while at rest can trigger contractions and movements of tiny muscle groups on the opposite side of the body (Penfield & Jasper, 1954; Penfield & Rasmussen, 1950; Rothwell et al. 1987). Conversely, however, activation of one half of the body can induce bilateral activation of the motor area.
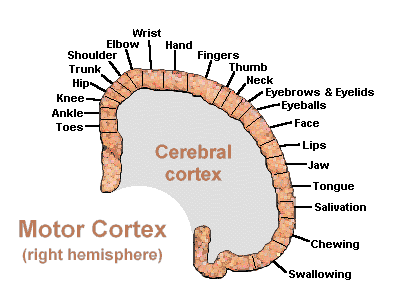
The primary motor area, however, is organized in a dual fashion in regard to the control over the body musculature. There is a one to one correspondence between neurons located in the primary motor area and specific muscles, which in turn makes fine motor functioning possible (Penfield & Jasper, 1954; Penfield & Rasmussen, 1950). However, there are also convergent connections as well, such that a single neuron innervates numerous muscle groups which in turn are innervated by yet other motor neurons (Humphrey, 2007; Shinoda, et al., 1981). In this manner, a neuron innervating a specific group of arm muscles, can also act on other muscles so as to coordinate movement.
Hence, a given muscle is also influenced by a number of motor neurons, such that a single muscle is represented by multiple neurons (Humphrey, 2007; Shinoda, et al., 1981). Multiple representation enables the motor cortex to coordinate a variety of movements, such that wide areas of the motor area become activated during even discrete movements of, for example a single finger (Colebatch et al., 1991; Sanes et al., 2013). Thus, activity is distributed over the motor area even when performing discrete movements.
Hence, the entire musculature of the body is neuronally represented in the motor cortex. Nevertheless, since certain muscle groups play a proportionately greater role in the performance of complex (vs simple) movements, a relatively greater number of neurons are involved in their representation. Thus, the hand and fingers have extensive cortical representation (Boroojerdik et al., 2011; Penfield & Jasper, 1954; Penfield & Rasmussen, 1950; Rothwell et al. 1987) whereas a smaller neuronal field is concerned with the elbow. The motor homunculus, therefore, is quite distorted.
Specifically, represented deep in the inferior medial portion of area 4 and moving anterior toward the superior medial frontal lobe are the toes, ankle, knee, hip, anal sphincter and genitals respectively. Circling up and over the superior surface to the lateral convexity are the shoulder, elbow, wrist, and hand, with extensive areas of neocortex devoted to the fingers and thumb, followed by the brow, eyelids, larynx, lips, jaw, and tongue which is located at the most inferior opercular portion of the precentral gyrus. Like the somesthetic neocortex which maintains a double representation of the body surface (e.g. a double body image) the primary motor area contains multiple representations of the body's musculature (Colebatch et al., 1991; Humphrey, 2007; Sanes et al., 2013; Shinoda, et al., 1981).
As noted, based on single cell recording, activity begins in the primary motor area following activation of the premotor and SMA and becomes maximally active at the time of movement. However, as based on single cell recording and functional imaging, neurons in the primary motor area also become active just prior to movement, increase their activity during movement, and will become increasingly active in response to direction, velocity, and the amount of force required to perform the movement (Ashe & Gerogopoulos, 2014; Crutcher & Alexander, 1990; Passingham, 2007). Indeed, this has also been demonstrated with evoked potentials. Activity will begin in the primary motor area prior to movement, and will increase as the time of movement onset approaches, and increases in amplitude during the course of movement (Ikeda & Shibasaki, 1992). In addition, these neurons will increase their activity depending on the amount of force involved in the movement, and in reaction to the direction and velocity of movement (Ashe & Georgopoulus, 2014).
Moreover, the motor area is exceedingly plastic and as indicated above, is capable of learning and memorizing and remembering (e.g. Carpenter et al., 2011). If a single muscle is repeatedly activated and/or as an individual becomes increasingly skilled at performing a certain motor tasks, the cortical territory representing those muscles will significantly increase (Pascual-Leone et al., 2014).
Paralysis.
Damage to the motor areas, or to the descending cortico-spinal tract initially results in a flaccid hemiplegia such that the muscles are completely without tone contralateral to the lesion (Adams & Victor, 2014; Brodal, 1981). If the examiner were to raise and release an effected arm, it would drop in a limp rag doll fashion.
Over the course of the next several days the muscles develop increased tone and there is resistance to passive movements. Reflexes become very brisk and spasticity and hyperreflexia are manifest. With massive lesions extending into the medial regions (where the leg is represented), the leg will become permanently extended and the arm will assume a flexed position (Adams & Victor, 2014; Brodal, 1981). After several weeks or months a very limited capacity to perform gross movements reappears. Fine movements are usually permanently lost (Brodal, 1981).
Exner's area is dependent on Broca's expressive speech area with which it maintains extensive interconnections. In fact, Exner's writing center extends to and appears to become coextensive with Broca's area (Lesser et al. 1984)--which in turn was originally a hand area--at least in primates. Broca's area possibly acts to organize and relay impulses received from the posterior language zones to Exner's area in instances where written expression is desired. Exner's area, in turn, transfers this information to the secondary and primary motor areas for final expression.
Electrical stimulation of this vicinity in awake moving patients has resulted in the arrest of ongoing motor acts, including the capacity to write or perfrom rapid alternating movements of the fingers (Lesser et al., 1984). In some instances, writing and speech arrest were noted.
Agraphia.
Lesions or seizure activity localized to this vicinity lead to deficiencies involving the elementary motoric aspects of writing; i.e. agraphia (Penfield & Roberts, 1959; Ritaccio et al. 1992; Tohgi et al. 2013). Grapheme formation becomes labored, incoordinated, and takes on a very sloppy appearance. Cursive handwriting is usually more disturbed than printing. In cases of well circumscribed lesions, usually there are no gross deficiencies of motor functioning or speech, although mild articulatory disturbances may be observed (e.g. lisping) as well as abnormalties involving fine motor control (Cf Lesser et al., 1984; Goodglass & Kaplan, 2000; Levine & Sweet, 1982.)
In cases of pure (frontal) agraphia, spelling may or may not be affected, whereas with parietal lesions spelling as well as writing is often abnormal. Rather, with left frontal lesions more frequently there are disturbances of grapheme selection such that the patient may seem to have "forgotten" how to form certain letters and/or may misplace or even add unnecessary letters when writing (Hecaen & Albert, 1978; Tohgi et al. 2013). When spelling orally or typing the ability to spell is often better preserved.
Damage localized to this vicinity can be secondary to perinatal trauma, tumors, or vascular abnormalities. Disturbances involving constructional or manipulospatial functioning are not apparent. In fact, one such patient whose damage was secondary to birth injury, although able to write or print only with great difficulty, was able to draw and paint with some professional acumen. However, his ability to copy letters was severely effected. Hence, disturbances secondary to lesions localized to Exner's area limited to abnormalities involving linguistic-symbolic grapheme motor control.
Ex=Exner's Writing Area; FEF=Frontal Eye Fields
The FEF receives projections from the primary and association visual cortices in the occipital lobe (17, 18, 19), the auditory association (22) and multimodal visual association areas (20) in the temporal lobe (Barbas & Mesulam, 1981; Jones et al., 1978; Jones & Powell, 1970), and the somatosensory association area (Crowne, 2003). It also shares interconnections with the caudate, superior colliculus, and oculomotor nucleus (Astruc, 1971; Knuzle & Akert, 1977; Segraves & Goldberg, 1987). Hence, the FEF receives information concerning the auditory, tactual, and visual environment and is multimodally responsive.
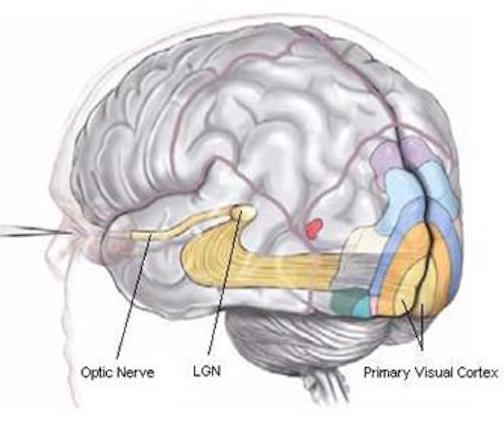
The FEF coordinates and maintaines eye and head movements, gaze shifts, and thus orienting and attentional reactions in response to predominantly visual, but also tactile and auditory stimuli (Barbas & Dubrovsky, 1981; Braun et al., 2006; Denny-Brown, 1966; Gottlieb et al. 2014; Latto & Cowey, 1971ab; MacAvoy, et al., 1991; Pragay et al. 1987; Segraves & Goldberg, 1987; Wagman et al. 1961). It is also involved in focusing attention on certain regions within the visual field, particularly the fovea (Wurtz et al. 1980; Segraves & Goldberg, 1987) as well as in making smooth pursuit movements (Gottlieb et al. 2014) and perhaps guiding eye movements while reading and when writing (e.g. Ritaccio et al. 1992).
In addition to visual tracking and supporting focused visual attention, neurons in the FEF demonstrate anticipatory activity (Gottlieb et al. 2014; Pragay et al. 1987); that is, firing before a response is made. In fact, these neurons will continue to fire at a high rate until the behavior is initiated. Yet other cells begin to fire only when the waiting period becomes prolonged (Pragay et al., 1987). In this regard, neurons in the FEF probably exert a countering influence so that attention does not drift and attention and eye position remains stabilized and focused on the primary retinal target.
Electrical stimulation of the FEF results in complete saccades of the eyes (Barbas & Dubrovsky, 1981; Wagman et al., 1981), as well as pupillary dilation. Moreover, cells in the FEF will fire slectively in response to stationary and moving stimuli, to objects which are within arms reach, as well as to tactual stimuli applied to the hands and/or mouth (Rizzolatti, Scandolara, Matelli, & Gentilucci, 1981ab). In fact, as an object approaches the face and mouth, some of these cells correspondingly increase their rate of activitity (Rizzolatti et al., 1981ab). Hence, cells within the FEF are highly involved in mediating sustained attention and orienting reactions of the head and eyes, maintaining visual fixation and modulating visual scanning, as well as coordinating eye-hand and hand to mouth as well as smooth pursuit eye movements.
Visual Scanning Deficits and Neglect.
Damage to the FEF can cause abnormalities in fixation, decreased sensitivity to stimuli throughout the visual field, slowed visual scanning and searching (Latto & Cowey, 1971ab; Teuber, 1964), inattention and neglect, as well as mislocation of sounds (Denny-Brown, 1966; Welch & Stuteville, 1958). Moreover, some subgroups of "schizophrenics" have been shown to suffer from smooth pursuit and sacadic abnormalities, which, based on neuropsychological tests, have been interpreted as indicative of FEF disturbances (O'Discoll, et al. 2013; Rosse et al. 1993).
With massive lessions, searching and responsiveness become so profoundly reduced that a complete unilateral neglect and failure to attend to any and all stimuli falling to one side of the body results (Heilman & Valenstein, 1972). Like confabulation, neglect is more frequently seen after right cerebral damage (chapter 10).
Even with less severe destruction performance on tasks involving Visual Search is disruped (Teuber, 1964) as is attention to visual detail (Luria, 1980). If shown and asked to describe a complex picture, patients may focus on only one detail, neglecting the remainder of the array, or look about haphazardly (Luria, 1980). It is probable that individuals with certain types of dyslexia suffer from similar abnormalities involving visual search and synthesis.
With large lesions (particularly when involving the right frontal area), they may tend to make leaps of judgment and impulsively guess and describe the meaning of the whole based on the perception of a fragment. For example, focusing only on a "drummer boy" in a battle scene they may describe the picture as being about "musicians" or "a rock band". Hence, such patients may erroneously extrapolate from an isolated detail around which they construct and confabulate a conclusion (Joseph, 2007a).
In the extreme, rather than a true analysis they may produce irrelevant associations as they not only have difficulty analyzing and synthesizing the different components of a visual array (Luria, 1980) but in correcting their impressions via seach and feedback. In these instances the lesion typically extends well beyond the confineds of the FEF.
With extensive damage involving the FEF and convexity, not only confabulation but Capgras syndrome (false identification) and reduplicative paramnesia have been reported--particularly if the damage is bilateral (Alexander, Stuss, & Benson, 1979; Benson, Gardner, & Meadows, 1979; Hecaen, 1964).
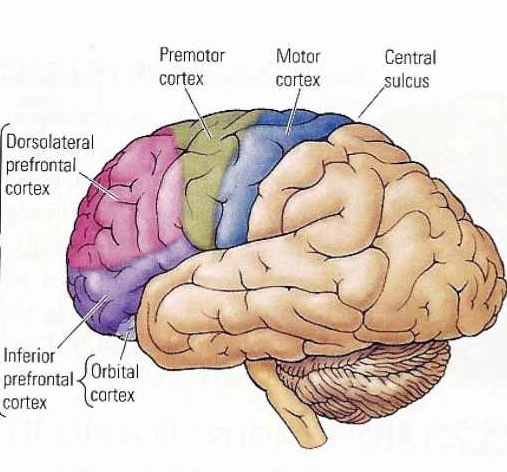







DVD - Brain Lectures
Six Lectures
3 DVD set = $29.95(+ shipping)
Pay By Paypal.com
DVD 1: Brain Overview
DVD 2: The Left Hemisphere, Brainstem, Midbrain, Thalamus
DVD 3: The Frontal Lobes: Frontal Lobotomy, Memory, Aphasia, Paralysis
DVD 4: The Parietal Lobes: Touch, Body-in-Space, Body Image, Hemi-Neglect, Phantom Limbs,
DVD 5: The Temporal Lobes: Language, Memory, Aphasia, Hallucinations, Face Recognition
DVD 6: The Limbic System: Amygdala, Hippocampus, Hypothalamus, Sex, Emotion, Memory, Stress, PTSD, Hallucinations