





Rhawn Gabriel Joseph, Ph.D.
BrainMind.com
Rnawn Joseph, Ph.D.
THE BRAIN RESEARCH LABORATORY
I. The Evolution of the Olfactory/Limbic System
II. Hypothalamus
A. Lateral & Ventromedial Hypothalamic nuclei
B. Hunger & Thrist
C. Pleasure & Reward
D. Emotion Incontinence: Laughter & Rage
III. Psychological Manifestations of Hypothalamic Activity
IV. Amygdala
A. Medial & Lateral Amygdala Nuclei
B. The Amygdala & Hypothalamus
C. Fear, Rage, Aggression
D. Social-Emotional Agnosia
V. Septal Nuclei
A. Rage & Quiescence
B. Contact Comfort & Septal Social Functioning
VI. Emotional Attachment & Amygdala-Septal Interactions
A. Limbic Abnormalities in Love & Socialization Skills
VII. The Cingulate Gyrus
A. The Evolution of Maternal Care
VIII. Limbic Language
A. Limbic Localization of Emotional Sound Production
B. Limbic Language & Mother-Infant Vocalization
IX. Sexual Differentiation of the Hypothalamus & Amygdala
A. Sex Differences in Language & Cognition
B. Sex Differences in Emotion
X. Hippocampus
A. Arousal, Attention & Inhibition
B. Learning & Memory
C. Hippocampal & Amygdala Interactions: Memory
D. Visual & Verbal Memory
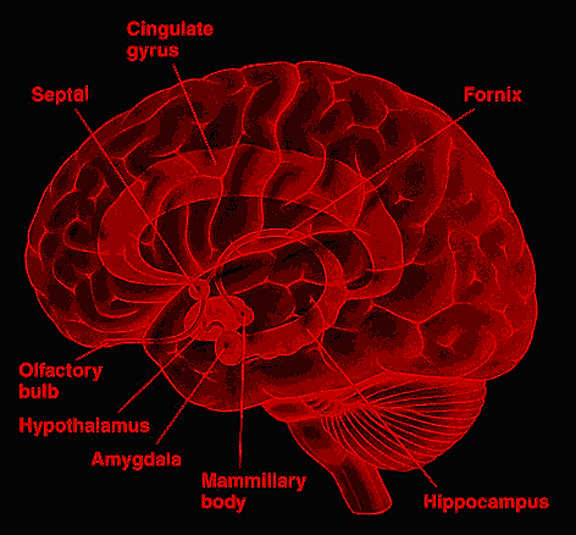
THE LIMBIC SYSTEM is buried within the depths of the cerebrum and consists of a collection of ancient brain structures which are preeminent in the mediation and expression of emotional, motivational, sexual, and social behavior. The limbic system is involved in learning and the formation of new memories, monitors internal homeostasis and basic needs such as hunger and thirst, controls the secretion of hormones involved in pregnancy and reactions to stress, and even makes possible the ability to experience orgasm, depression, fear, rage, and love.
Broadly, these limbic system nuclei include the hypothalamus, amygdala, hippocampus, septal nuclei, and cingulate gyrus. Also related to limbic system functioning are portions of the reticular activating system, the orbital frontal and inferior temporal lobes, as well as parts of the thalamus and cerebellum. Indeed, the limbic system is not only exceedingly ancient but originally provided the foundation for the development and evolution of much of the brain.
I. THE EVOLUTION OF THE OLFACTORY/LIMBIC SYSTEM
About 1 billion years ago a cellular metamorphosis of paramount importance resulted in the creation of a completely unique type of cell, the neuron. These nerve cells in turn were especially responsive to light as well as chemical (olfactory and pheromonal) messages. Over the course of evolution, as the number of secreting and transmitting nerve cells that a creatures possessed increased, a network of interlinked neurons, called the "nerve net" was soon fashioned. Soon tiny neural ganglia, composed of colonies of similarly functioning nerve cells began to form in the anterior head region of various ancient and primitive creatures.
By time the first vertebrates and fish begin to swim the oceans, around 500 million years ago, the first primitive lobes of the brain had also become fashioned through the collectivization of these neural ganglia. This included the olfactory-limbic lobe (the forebrain), which was concerned with the detection of olfactory/pheromonal chemicals that might betray the presence of a predators, prey, or a mate; the optic lobe/tectum of the midbrain which was responsive to visual messages, and the hindbrain which was concerned with movement. By 450,000 years ago, the first sharks had acquired a limbic system, which they, like modern humans, still possess today.
II. HYPOTHALAMUS
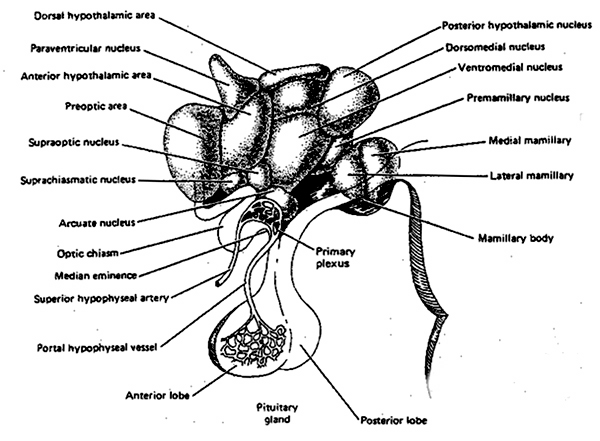
The hypothalamus is an exceedingly ancient structure and unlike most other brain regions it has maintained a striking similarity in structure throughout phylogeny and apparently over the course of evolution. The hypothalamus is fully functional at birth and is highly involved in all aspects of endocrine, hormonal, visceral and autonomic functions and mediates or exerts controlling influences on eating, drinking, the experience of pleasure, rage, and aversion. The hypothalamus is the central core from which all emotions derive their motive force.
The hypothalamus is also sexually differentiated. That is, structurally and functionally the hypothalamus of men and women are sexually dissimilar.
A. LATERAL & VENTROMEDIAL HYPOTHALAMIC NUCLEI
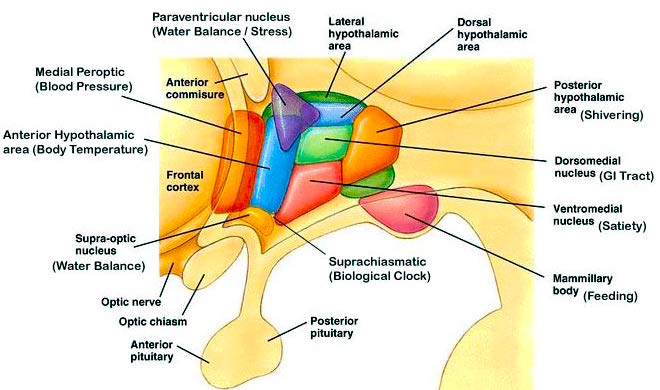
Although the hypothalamus consists of several distinct regions and subgroups, the lateral and medial (ventromedial) hypothalamic nuclei play particularly important roles in almost all aspects of emotion and internal homeostasis. They also appear to share a somewhat antagonistic relationship and act to exert counterbalancing influences on each other.
For example, the medial hypothalamus controls parasympathetic activities (e.g. reduction in heart rate, increased peripheral circulation) and exerts a dampening effect on certain forms of emotional/motivational arousal. The lateral hypothalamus mediates sympathetic activity (increasing heart rate, elevation of blood pressure) and is involved in controlling the metabolic and somatic correlates of heightened emotionality.
B. HUNGER & THIRST
The lateral and medial region are highly involved in monitoring internal needs such as hunger and thirst. For example, both nuclei contain receptors which are sensitive to the body's fat content (lipostatic receptors) and to circulating metabolites such as glucose, which together indicate the need for food and nourishment. The lateral hypothalamus also appears to contain osmoreceptors which determine if water intake should be altered. Both hypothalamic nuclei also become highly active immediately prior to and while the organism is eating or drinking.
For example, the lateral region alters it's activity when the subject is hungry and simply looking at food. If the lateral hypothalamus is electrically stimulated a compulsion to eat and drink results. Conversely, if the lateral area is destroyed there results aphagia and adipsia so severe animals will die of starvation. If the medial hypothalamus is surgically destroyed, inhibitory influences on the lateral region appear to be abolished such that hypothalamic hyperphagia and severe obesity result.
Overall, it appears that the lateral hypothalamus is involved in the initiation of eating and acts to maintain a lower weight limit such that when the limit is reached the organism is stimulated to eat.
Conversely, the medial regions seems to be involved in setting a higher weight limit such that when these levels are approached it triggers the cessation of eating. That is, the medial area seems to act as a satiety center; but, a center that can be overridden. In part, these nuclei exert these differential influences on eating and drinking via motivational/emotional influences they exert on other brain nuclei (e.g. via reward or punishment).
C. PLEASURE & REWARD.
In 1952, R.G. Heath reported what was then considered remarkable. Electrical stimulation near the septal nuclei elicited feelings of pleasure in human subjects: "I have a glowing feeling. I feel good!" Subsequently, in 1954 James Olds and Peter Milner reported that rats would tirelessly press a lever in order to receive electrical stimulation via tiny electrodes planted in this same region. Olds and Milner, in fact, concluded that stimulation in this region of the brain "has an effect which is apparently equivalent to that of a conventional primary reward." Even hungry animals would demonstrate a preference for self-stimulation over food.
Feelings of pleasure have since been obtained following electrical excitation to a number of diverse limbic areas including the olfactory bulbs, amygdala, hippocampus, cingulate gyrus, the basal ganglia, thalamus, reticular formation, medial forebrain bundle, and orbital frontal lobes. However, the greatest area of concentration and the highest rates of self-stimulatory activity were found to occur in the lateral hypothalamus. Indeed, according to Olds, animals "would continue to stimulate as rapidly as possible until physical fatigue forced them to slow or to sleep."
More recently, the lateral hypothalamus (as well as the amygdala and other limbic nuclei) have been found to have nerve cells which produce and are responsive to opiate-like substances, i.e. enkephalins. Hence, when an individual is injected to with various narcotic substances, it is these limbic nuclei which respond with feelings of pleasure.
In contrast to the lateral hypothalamus and it's involvement in pleasurable self-stimulation, activation of the medial hypothalamus is so aversive that subjects will work to reduce it --apparently so as to obtain relief (e.g. active avoidance).
In this regard, when considering behavior such as eating, it might be postulated that when upper weight limits (or nutritional requirements) are met, the medial hypothalamus becomes activated which in turn leads to behavior which terminates it's activation (e.g. cessation of eating). In fact, it is probably in this manner that the hypothalamus is able to exert considerable influence on a variety of behaviors, either acting to reward one's actions, or to generate feelings of aversion so that one is less likely to act in a similar manner in the future.
D. EMOTIONAL INCONTINENCE: LAUGHTER & RAGE
Although highly involved in all aspects of emotion and motivational functioning, the emotional states elicited by the hypothalamus are very primitive, diffuse, undirected and unrefined, being limited to pleasure in general, or aversion/unpleasure in general. It is for this reason that ancient and primitive animals are also very limited in their ability to express and perceive emotion. Higher order emotional reactions (e.g. desire, love, hate, etc.) require the involvement of other limbic regions as well as the participation of the more recently evolved regions of the brain, the neocortex (i.e. "new brain").
Nevertheless, due to its involvement in the generation of positive and negative emotions, not surprisingly, when the hypothalamus has been injured or is made to function abnormally, extremely positive or negative reactions can also be elicited, including rage and uncontrolled laughter. For example, laughter has been noted to occur with hilarious or obscene speech--usually as a prelude to stupor or death--in cases where tumor has infiltrated the hypothalamus. In several instances it has been reported that in the course of neurosurgery involving the hypothalamus, patients "became lively, talkative, joking, and whistling each time the hypothalamus was manipulated."
In one case, the patient became excited and began to sing. Some individuals with hypothalamic damage have in fact died laughing. However, such patients claim that their laughter does not reflect there true feelings. Hence, laughter in these instances has been referred to as "sham mirth." Moreover, the type of emotional reaction elicited is dependent on which region of the hypothalamus has been injured or activated.
Stimulation of the lateral hypothalamus, for example, can induce extremes in emotionality, including intense attacks of rage accompanied by biting and attack upon any moving object. If this nucleus is destroyed, aggressive and attack behavior is abolished.
Hence, the lateral hypothalamus is responsible for rage and aggressive behavior, including attack and predatory actions, which coincides with its involvement with eating. In contrast, stimulation of the medial region counters the lateral hypothalamus such that rage reactions are reduced or eliminated. If the medial region is destroyed there results lateral hypothalamic release and the triggering of extreme savagery.
Nevertheless, like "sham mirth", rage reactions elicited in response to direct electrical activation of the hypothalamus immediately and completely dissipate when the stimulation is removed. As such, these outbursts have been referred to as "sham rage".
III. PSYCHOLOGICAL MANIFESTATIONS OF HYPOTHALAMIC ACTIVITY
Phylogenetically and from an evolutionary perspective, the appearance and development of the hypothalamus predates the emergence and differentiation of all other limbic nuclei, e.g., amygdala, septal nucleus, hippocampus. It constitutes the most primitive, archaic, reflexive, and purely biological aspect of the psyche.
Biologically the hypothalamus serves the body tissues by attempting to maintain internal homeostasis and by providing for the immediate discharge of tensions in an almost reflexive manner. Hence, as based on studies of lateral and medial hypothalamic functioning, it appears to act reflexively, in an almost on/off manner so as to seek or maintain the experience of pleasure and escape or avoid unpleasant, noxious conditions.
Emotions elicited by the hypothalamus are largely undirected, short-lived, and unconnected with events occurring within the external environment, being triggered reflexively and without concern or understanding regarding consequences. Direct contact with the real world is quite limited and almost entirely indirect as the hypothalamus is largely concerned with the internal environment of the organism. It has no sense of morals, danger, values, logic, etc., and cannot feel or express love or hate. Although quite powerful, hypothalamic emotions are largely undifferentiated, consisting of feelings such as pleasure, unpleasure, aversion, rage, hunger, thirst, etc.
As the hypothalamus is concerned with the internal environment much of it's activity occurs outside conscious-awareness. Moreover, being involved in maintaining internal homeostasis, via, for example, it's ability to reward or punish the organism with feelings of pleasure or aversion, it tends to serve what Sigmund Freud described as the "pleasure principle".
IV. AMYGDALA
In contrast to the primitive hypothalamus, the amygdala is preeminent in the control and mediation of all higher order emotional and motivational activities, including the capacity to form emotional attachments and to feel love. Neurons located in the amygdala are able to monitor and abstract from the sensory array stimuli that are of motivational significance so as to organize and express appropriate feelings and behaviors. This includes the ability to discern and express even subtle social-emotional nuances such as friendliness, fear, affection, distrust, anger, etc., and at a more basic level, determine if something might be good to eat. In fact, amygdaloid neurons respond selectively to the flavor of certain preferred foods, as well as to the sight or sound of something that might be especially desirable to eat.
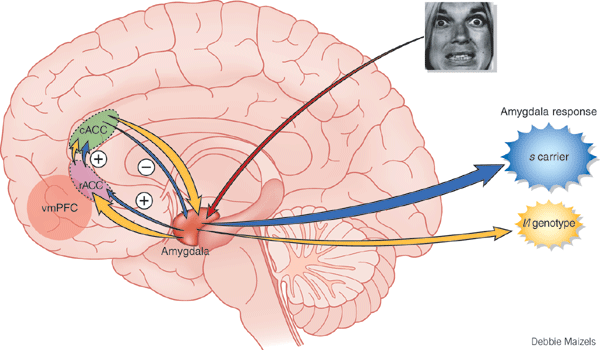
Moreover, some neurons located in the amygdala are responsive to faces and facial emotions conveyed by others. Many neurons are also able to respond to visual, tactual, olfactory, and auditory stimuli simultaneously. Hence, many amygdaloid neurons are predominantly polymodal, responding to a variety of stimuli from different modalities. It is in this manner that the amygdala has come to be involved not only in emotion, but attention, and learning and memory, for multimodal assimilation of various sensory impressions occurs in this region.
A. MEDIAL & LATERAL AMYGDALA NUCLEI
The amygdala is buried within the depths of the anterior-inferior temporal lobe and consists of two major nuclear groups. These are a phylogenetically ancient anteromedial group (or medial amygdala) which is involved in olfaction, pheromonal perception, and motor activity (via rich interconnections with the basal ganglia), and a relatively newer basolateral division (lateral amygdala) which is maximally developed among humans.
Like the lateral and medial hypothalamus, the medial and lateral amygdala are rich in opiate receptors and cells containing enkephalins and both subserve different functions. For example, the medial amygdala is highly involved in motor, olfactory and sexual functioning, whereas the lateral division is intimately involved in all aspects of higher order emotional activity including the generation of selective attention. That is, the amygdala acts to perform environmental surveillance and can trigger orienting responses as well as mediate the maintenance of attention if something of interest or importance were to appear. Hence, electrical stimulation of the lateral division can initiate quick and/or anxious glancing and searching movements of the eyes and head such that the organism appears aroused and highly alert as if in expectation of something that is going to happen.
Indeed, via its rich interconnections with the inferior, middle, and superior temporal lobes, as well as other neocortical regions, the lateral amygdala is able to sample and influence the auditory, somesthetic, and visual information being received and processed in these areas, as well as scrutinize this information for motivational and emotional significance. It is through the lateral division that emotional meaning and significance can be assigned to as well as extracted from that which is experienced.
B. THE AMYGDALA & HYPOTHALAMUS
The amygdala, overall, maintains a functionally interdependent relationship with the hypothalamus. It is able to modulate and even control rudimentary emotional forces governed by the hypothalamic nucleus. However, it also acts as the behest of hypothalamically induced drives. For example, if certain nutritional requirements need to be meet, the hypothalamus signals the amygdala which then surveys the external environment for something good to eat. On the other hand, if the amygdala via environmental surveillance were to discover a potentially threatening stimulus such as a predator, it acts to excite and drive the hypothalamus as well as the motor centers, so that the organism is mobilized to take appropriate action.
When the hypothalamus is activated by the amygdala, instead of responding in an on/off manner, cellular activity continues for an appreciably longer time period. The amygdala can tap into the reservoir of emotional energy mediated by the hypothalamus so that certain ends may be attained.
C. FEAR, RAGE & AGGRESSION
Initially, electrical stimulation of the amygdala produces sustained attention and orienting reactions. If the stimulation continues fear and/or rage reactions are elicited. When fear follows the attention response, the pupils dilate and the subject will cringe, withdraw, and cower. This cowering reaction in turn may give way to extreme fear and/or panic such that the animal will attempt to take flight.
Among humans, the fear response is one of the most common manifestations of amygdaloid electrical stimulation. Moreover, unlike hypothalamic on/off emotional reactions, attention and fear reactions can last up to several minutes after the stimulation is withdrawn. In addition to behavioral manifestations of heightened emotionality, amygdaloid stimulation can also result in intense changes in emotional facial expression. This includes facial contortions, baring of the teeth, dilation of the pupils, widening or narrowing of the eye-lids, flaring of the nostrils, tearing, as well as sniffing, licking, and chewing. In fact, epileptic seizure activity in this area (i.e. temporal lobe epilepsy) often induces involuntary chewing, and smacking of the lips and licking.
In many instances, rather than fear, stimulation of the amygdala results in anger, irritation, and rage which seems to gradually build up until finally the animal or human will attack. Unlike hypothalamic "sham rage", amygdaloid activation results in attacks directed at something real, or, in the absence of an actual stimulus, at something imaginary. Moreover, rage and attack will persist well beyond the termination of the electrical stimulation of the amygdala.
In fact, the amygdala remains electrophysiologically active for long time periods even after a stimulus has been removed (be it external-perceptual, or internal-electrical) such that is appears to continue to process--in the abstract--information even when that information is no longer observable. Moreover, tumors in this area can trigger violent rage attacks. A famous example of this is Charles Whitman, who in 1966 climbed a tower at the University of Texas carrying a high powered hunting rifle and for the next 90 minutes shot at everything that moved, killing 14, wounding 38. Post-mortem autopsy of his brain revealed a glioblastoma multiforme tumor the size of a walnut compressing the amygdaloid nucleus.
D. SOCIAL-EMOTIONAL AGNOSIA
Among primates and mammals, bilateral destruction of the amygdala significantly disturbs the ability to determine and identify the motivational and emotional significance of externally occurring events, to discern social-emotional nuances conveyed by others, or to select what behavior is appropriate given a specific social context. Bilateral destruction of both amygdalas (located in the right and left temporal lobe) usually results in increased tameness, docility, and reduced aggressiveness in cats, monkeys and other animals and humans. It also lowers responsiveness to aversive and social stimuli, and reduces fearfulness, competitiveness, dominance, and social interest. Indeed, this condition is so pervasive that subjects seem to have tremendous difficulty discerning the meaning or recognizing the significance of even common objects -- a condition sometimes referred to as "psychic blindness", or, the "Kluver-Bucy syndrome". However, it is important to note that although Drs. Kluver and Bucy reported this in 1937, this condition had first been reported in 1888 by Drs. Brown and Shaefer.
Like an infant (who similarly is without a fully functional amygdala), individuals with bilateral amygdala destruction engage in extreme orality and will indiscriminately pick up various objects and place them in their mouth regardless of its appropriateness. There is a repetitive quality to this behavior, for once they put it down they seem to have forgotten that they had just explored it, and will immediately pick it up and place it again in their mouth as if it were a completely unfamiliar object.
Hence, humans as well as animals with bilateral amygdala destruction, although able to see and interact with their environment, respond in an emotionally blunted manner, and seem unable to recognize what they see, feel, and experience. Things seem stripped of meaning. This condition pervades all aspects of higher level social-emotional functioning including the ability to appropriately interact with loved ones. As might be expected, maternal behavior is severely affected. According to Dr. A. Kling, mothers will behave as if their "infant were a strange object be be mouthed, bitten and tossed around as though it were a rubber ball".
Among primates who have undergone bilateral amygdaloid removal, once they are released from captivity and allowed to return to their social group, a social-emotional agnosia becomes readily apparent as they no longer respond to or seem able to appreciate or understand emotional or social nuances. Indeed, they appear to have little or no interest in social activity and persistently attempt to avoid contact with others. If approached they withdraw, and if followed they flee. Indeed, they behave as if they have no understanding of what is expected of them or what others intend or are attempting to convey, even when the behavior is quite friendly and concerned. Among adults with bilateral lesions, total isolation seems to be preferred.
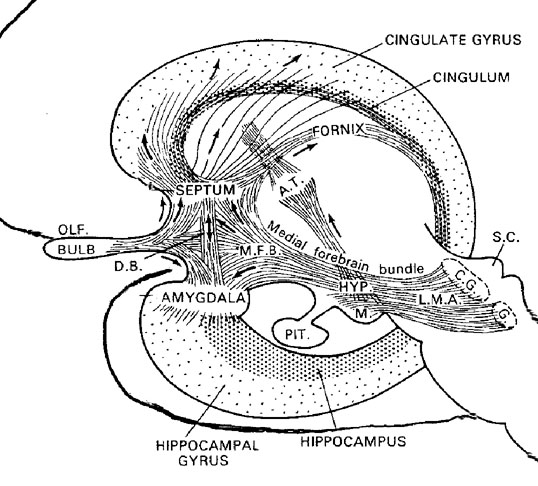
It is thus apparent that the amygdala, in conjunction with other limbic tissue, such as septal nuclei and the more recently evolved transitional limbic cortex, the cingulate gyrus, is highly involved in all aspects of social and emotional functioning. In fact, as argued by Dr. R. Joseph in a number of articles, it appears that the differential maturation of these limbic structures, in particular, that of the amygdala, septal nuclei, and cingulate gyrus, are responsible for seeking contact comfort and forming of emotional and loving attachments during infancy.
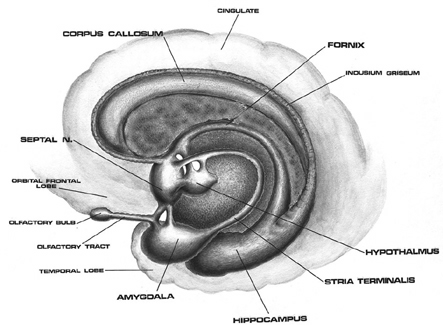
V. SEPTAL NUCLEI
The septal nuclei, like the amygdala is very ancient, and appears to develop out of the hypothalamus. Phylogenetically and presumably, ontogentically, it seems to mature following the development of the amygdala, but at about the same time as the hippocampus, a limbic system structure involved in the formation of memory. The septal nuclei also increases in relative size and complexity as we ascend the ancestral tree, attaining its greatest degree of development in humans.
The septal nuclei lies in the medial portions of the hemispheres, just anterior to the hypothalamus, and maintains rich interconnections with all regions of the limbic system. Unfortunately, unlike other limbic tissue, the functioning of the septal nuclei is still not well understood. Nevertheless, it appears to maintain a complementary relationship with the hippocampus, but an oppositional, and sometimes antagonistic relationship with the amygdala. For example, the amygdala appears to act so as to either facilitate or inhibit septal functioning whereas septal influences on the amygdala are largely inhibitory. However, in large part, the amygdala and septal nuclei appear to exert the majority of their counterbalancing influences on the emotional functioning of the hypothalamus with which they both maintain rich interconnections.
A. RAGE & QUIESCENCE
A primary activity of the septal nucleus appears to be that of reducing extremes in emotionality and arousal, and maintaining the organism in state of quiescence and readiness to respond. Stimulation of the septum acts to reduce blood pressure and heart rate, induces adrenocortical secretion, counters lateral hypothalamic self-stimulatory activity, inhibits aggressive behavior and suppresses the expression of rage reactions following hypothalamic stimulation.
If the septal nucleus is destroyed, these counterbalancing influences are removed such that initially there results dramatic increases in aggressive behavior. In fact, bilateral lesions of the septal nuclei can trigger explosive emotional reactivity to tactile, visual, or auditory stimulation such that the animal may attempt to attack or run away. However, if the amygdala is subsequently destroyed, the septal rage and emotional reactivity are completely attenuated. However, when the amygdala remains intact, septal lesions appear to result in a loss of modulatory and inhibitory restraint.
B. CONTACT COMFORT & SEPTAL SOCIAL FUNCTIONING
Although initially destruction of the septal nuclei results in rage reactions, within a few weeks this aggressiveness subsides and/or completely disappears. However, a generalized tendency to over respond and a generalized failure to inhibit emotional responsiveness persists, and animals so effected tend to demonstrate an extreme and indiscriminate need for social and physical contact. That is, in contrast to amygdaloid lesions which produce a severe social-emotional agnosia and social avoidance, septal lesions produce a dramatic and persistent increase in social cohesiveness.
These findings suggest that the normal, intact amygdala appears to promote social behavior whereas the septal nucleus seems to counter socializing tendencies. Hence, with destruction of the septal nuclei (which results in a release of the amygdala), the drive for social contact appears to be irresistible such that persistent attempts to make physical contact occurs--even with species quite unlike their own.
For example, septal lesioned rats, unlike normals, will readily seek out mice (to which they are normally indifferent) or rabbits (which they usually avoid). If presented with a choice of an empty (safe) chamber or one containing a cat, septal lesioned rats persistently attempt to huddle and crawl upon this normally feared creature, even when the cat is acting perturbed. If a group of septally lesioned animals are placed together, extreme huddling results.
So intense is this need for contact comfort following septal lesions, that if other animals are not available they will seek out blocks of wood, old rags, bare wire frames or walls.
Among humans with right sided or bilateral disturbances in septal functioning (such as due to seizure activity being generated in this region), a behavior referred to as "stickiness" is sometimes observed. Such individuals seek to make repeated, prolonged, and often inappropriate contact with anyone who is available or who happens to be near by so as to tell them stories, jokes or merely pass the time. Moreover, they refuse to take a "hint," and do not depart unless given a direct request to do so.
VI. EMOTIONAL ATTACHMENT & AMYGDALA-SEPTAL INTERACTIONS
Physical, social and emotional interaction and contact during infancy is critically important to the child's well being as well as his or her neurological, sensory, cognitive, intellectual, social and emotional development. Indeed, babies need their "mamas" and all the love and attendant physical and emotional interaction they can get. The more an infant is held, stroked and spoken to, and the greater the visual divergence of his surroundings, the greater will be its resilience and capability to adapt to negative emotional and physical onslaughts and to withstand stressful extremes later in life. In fact, the very cells of the nervous system will prosper by growing larger and more complex.
So great is the need for stimulation that until 6-7 months of age most children will eagerly and indiscriminately seek social and physical contact from anyone including complete strangers. Indiscriminate social interaction is not merely a manifestation of friendliness but serves a specific purpose: it maximizes opportunities for social and physical contact and interaction. Like hunger and the desire for food (which is mediated by the hypothalamus) there is a physical drive and hunger for social, emotional, and physical stimulation (which is mediated by the amygdala).
At about 7 months of age the infant becomes more discriminate in his or her interactions and it is during this time period that a very real and specific attachment (e.g. to one's mother) becomes progressively more intense and stable. This does not mean that prior to this period the mother is not highly important to the infant, but rather maximal social interaction takes precedence during the first critical months of life. In other words, the baby needs more contact than a single mother is capable of providing.
After these specific attachments such as to mother have been formed, most children increasingly begin to show anxiety, fear and even flight reactions at the approach of a stranger. By one year of age 90% of children respond aversively to strangers. This also serves a purpose for it maximizes the bond with mother and insures that a child who can crawl and maneuver through space does not indiscriminately attach to and wander off with a stranger.
Thus the infants initial seeking of indiscriminate social contact is followed at a later age by progressively narrowed contact seeking. According to a theory developed by Dr. R. Joseph, these stages of emotional development coincide with the maturation of different nuclei in the limbic system of the brain; the amygdala, septal nuclei and cingulate gyrus.
As noted, the septal nucleus and amygdala often act in balanced opposition. That is, the septal nuclei appears to be highly involved in social and intimate contact seeking, but in a fashion quite different from the amygdala. The normal amygdala, which matures before the septal region promotes social contact seeking, whereas the normal, undamaged septal nuclei, which matures later, acts to inhibit and restrict these tendencies so that they are directed and focused (such as upon one person), rather than being generalized and indiscriminate. These two regions of the brain, in conjunction with the cingulate gyrus are highly interactive and crucially important in the formation of our first and earliest attachments, as well as those later in life.
It is these same limbic nuclei which later in life are involved in the ability to feel love (as well as hate and anger) for, and attachment to, a loved one. That is, the limbic system controls the basic aspects of emotion, such as love, hate, anger, rage, fear, pleasure, the desire to bond together, as well as biological drives, including hunger, thirst, and even the capacity to experience orgasm during sex. Often all these impulses and needs at one time or another becomes associated with mother or the primary caretaker, and later in life (to a considerable degree), with a spouse. Even the presence or absence of mother can at one time or another elicit these responses (e.g. rage when the infant is not being held or fed).
Similarly, due to limbic attachment, the rejecting actions of a mate elicit limbic reactions including infantile feelings of rage and abandonment. That is, the amygdala striving to maintain the bonds of love responds with rage when the bond is severed, and the hypothalamus, feeling likewise, responds similarly. Even the murderous desire to kill one's spouse can be elicited. Indeed, loss of love, such as occurs when a relationship ends, seconded only by jealousy and money are prime elicitors of such murderous feelings and are due to the high involvement of the limbic system in all affairs of the heart.
A. LIMBIC ABNORMALITIES IN LOVE & SOCIALIZATION SKILLS
If contact with others is restricted during the early phases of infant development, then the ability to interact successfully with others at a later stage of of life is retarded. That is, the infant and child must experience love and nurturance during this time period, otherwise these limbic nuclei will not develop and interact normally. If these interactional needs are not met during this critical period of development, gross abnormalities can result. Children will lose the ability to form emotional attachments with others, sometimes for the rest of their lives.
This is even true among non-human animals. Kittens which are not handled or stroked by humans soon become "wild" and unapproachable even when they have otherwise been exposed to people on a daily basis. Similarly, young children and infants who are separated from their parents and who fail to receive necessary loving and social stimulation are also affected adversely. They have difficulty forming emotional attachments and even their brains may not properly develop. If not adequately physically and emotionally stimulated, the child may even die.
In other words, if a child is not firmly attached to a mother figure and has been neglected early in life, the ability to form attachments increasingly narrows and then disappears, possibly forever. The child becomes attached to no one and its ability to form loving attachments later in life will be abnormal if drastic countermeasures are not taken. This is because cells in the amygdala, not receiving sufficient and appropriate stimulation begin to die and atrophy from disuse; just like a muscle if unused. "Use it or lose it." Once these limbic neurons die or if certain interconnections between different regions are not maintained, they are no longer able to respond appropriately to physical, emotional and social interaction.
VII. THE CINGULATE GYRUS A. THE EVOLUTION OF MATERNAL CARE
As noted, most creatures, including sharks, amphibians, reptiles and fish, possess a limbic system, consisting of an amygdala, hippocampus, hypothalamus, and septal nuclei. It is these limbic nuclei which enable a group of fish to congregate together, i.e. to school, or for reptiles (creatures who first began to roam the planet about 300 million years ago) to form territories and very loosely organized social aggregates consisting of an alpha male and female, several sub-females, and a few juveniles. Such creatures, however, although sometimes showing parental investment, generally do not provide long term care for their young and do not produce complex meaningful vocalizations, although, like amphibians they do produce very limited socially meaningful sounds, which in turn appear to be generated and perceived by limbic nuclei such as the amygdala.
Nevertheless, although in possession of a limbic system, reptiles, and other non-mammalian species are lacking the more recently acquired cingulate cortex which appears to have begun to evolve around 250 million years ago when reptiles diverged to form repto-mammals (the therapsids) who in turn evolved into mammals and then primates. It was with the appearance of the repto-mammals that the first evidence of suckling of infants and long term maternal care came into being. Indeed, it has been postulated by Paul Maclean (who in fact coined the term "limbic system") as well as by Dr. R. Joseph, that the cingulate (in conjunction with the amygdala and septal nuclei) is largely responsible for the appearance of maternal feelings, and the evolution of the family.
However, primates and other mammals, in addition to limbic and transitional limbic cingulate cortex, are also equipped with the six to seven layered neocortex which evolved approximately 100 million years ago and which covers the old brain like a shroud. However, like the amygdala, the cingulate has reached its maximal size among humans and maintains rich interconnections with the neocortex as well as with the older portions of the limbic system such as the amygdala and hippocampus.
Among humans and lower mammals, destruction of the anterior cingulate results a loss of fear, lack of maternal responsiveness and severe alterations in socially appropriate behavior. Humans will often become initially mute and socially unresponsive, and when they speak, their vocal melodic-inflectional patterns and the emotional sounds they produce sound abnormal.
Animals, such as monkeys who have suffered cingulate destruction will also become mute, will cease to groom or show acts of affection and will treat their fellow monkeys as if they were inanimate objects. For example, they may walk upon and over them as if they were part of the floor or some obstacle rather than a fellow being. In other words, their behavior is more typical of a reptile than a primate. Maternal behavior is also abolished following cingulate destruction, and the majority of infants soon die from lack of care.
More importantly, when the cingulate cortex is electrically stimulated, the separation cry, similar if not identical to that produced by an infant, is elicited. In fact, it appears that the cingulate, in conjunction with the amygdala and other limbic tissue are not only responsible for the development of long term infant care, but the initial production of what would become language. This has been referred to by Joseph as "limbic language." In fact, be it humans or reptiles the limbic system is preeminent in the mediation, production, and comprehension of emotional-social sounds, including sex differences in their production.
VIII. LIMBIC LANGUAGE
Phylogenetically and ontogenetically, the original impetus to vocalize springs forth from roots buried within the depths of the ancient "limbic lobe" a term coined by Papez in 1937. Although non-humans do not have the capacity to speak, they still vocalize, and these vocalizations are primarily limbic in origin being evoked in situations involving sexual arousal, terror, anger, flight, helplessness, and separation from the primary caretaker when young.
The first vocalizations of human infants are similarly emotional in origin and limbically mediated, consisting predominantly of sounds indicative of pleasure and displeasure. Indeed, these sounds and cries are produced soon after birth, indicating they are innate, and are produced even by infants born deaf and blind. Similarly, apes and monkeys reared in isolation or with surgically muted mothers also produce appropriately sounding complex emotional calls in order to convey a wealth of information, including the presence of danger. Moreover, they will respond to these same calls with appropriate reactions, even when they had never before been heard.
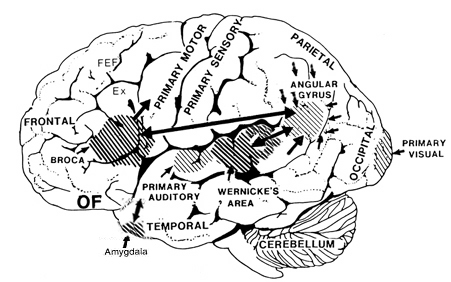
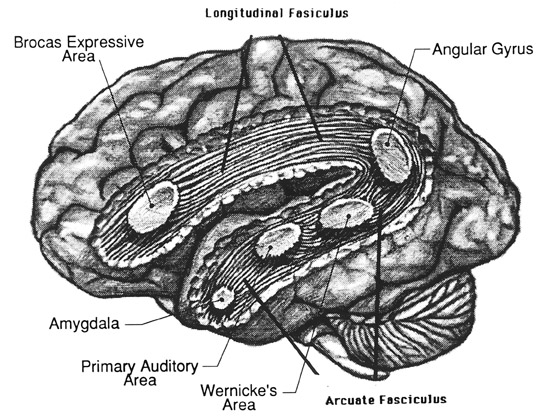
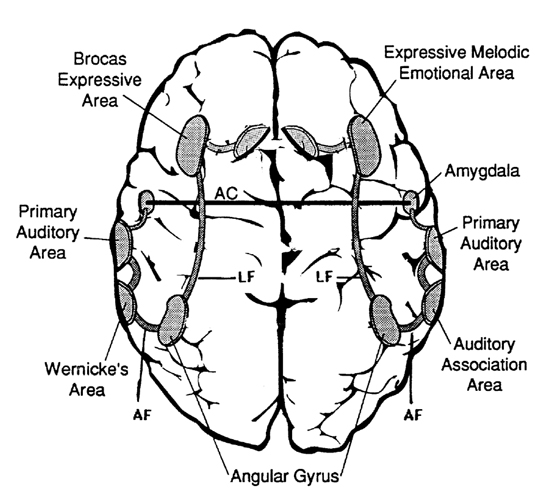
Emotional cries and warning calls have been produced via electrode stimulation of wide areas throughout the limbic system. Nevertheless, the type of cry elicited, in general, depends upon which limbic nuclei has been activated. For example, portions of the septal nuclei, hippocampus, and medial hypothalamus have been repeatedly shown to be generally involved in the generation of negative and unpleasant mood states, whereas the lateral hypothalamus and amygdala, and portions of the septal nuclei, are associated with pleasurable feelings.
Not surprisingly, areas associated with pleasurable sensations often give rise, when sufficiently stimulated, to pleasurable calls, whereas those linked to negative mood states, will trigger cries of alarm and shrieking. However, of all limbic nuclei, the amygdala and cingulate gyrus appear to be the most vocal.
In humans and animals a wide range of emotional sounds have been evoked through amygdala activation, including those indicative of pleasure, sadness, happiness, and anger. Conversely, in humans, destruction limited to the amygdala, the right amygdala in particular, has abolished the ability to sing, convey melodic information or to enunciate properly via vocal inflection and can result in great changes in pitch and the timbre of speech. Even the capacity to perceive and respond appropriately to social-emotional cues is abolished.
However, in the cingulate gyrus, completely different emotional calls can be elicited from electrodes which are immediately adjacent, and, the calls do not always correlate with the mood state. This suggests considerable flexibility within the cingulate which also appears to have the capability of producing emotional sounds that are not reflective of mood. This suggests a high degree of voluntary control within the cingulate. However, of the many sounds produced, the separation cry of the infant is one of the most significant, particularly in regard to the evolution of language. It is from the cingulate where the separation cry is most frequently elicited.
B. LIMBIC LANGUAGE & MOTHER-INFANT VOCALIZATION
Among social terrestrial vertebrates the production of sound is very important in regard to infant care, for if an infant becomes lost, separated, or in danger, a mother would have no way of quickly knowing this by smell alone. Such information would have to be conveyed via a cry of distress or a sound indicative of separation fear and anxiety. It would be the production of these sounds which would cause a mother to come running to the rescue. Hence, the first forms of limbic social-emotional communication was probably produced in a maternal context.
Indeed, considerable vocalizing typically occurs between human and non-human mammalian mothers and their infants, and the infants of many species, including primates, will often sing along or produced sounds in accompaniment to those produced by their mothers. In fact, among primates, females are more likely to vocalize and utter alarm calls when they are near their infants versus non-kin, and vice versa, and adult males are more likely to call or cry when in the presence of their mother or an adult female vs an adult male. Similarly, infant primates will loudly protest when separated from their mother so long as she is in view and will quickly cease to vocalize when isolated. It thus appears that the purpose of these vocalizations are to elicit a response from the mother.
Hence, the production of emotional sounds appears to be limbically linked and associated with maternal-infant care, and with interactions with an adult female. In fact, human females in general tend to vocalize more so than males and their speech tends to be perceived as friendlier and more social.
It is important to note, however, that the hypothalamus, septal nuclei, as well as the periquaductal gray (which is located in the midbrain) are also important components in the formulation of limbic language. Given the role of these limbic nuclei in sex related differences in cognition and behavior, it is perhaps highly likely that they may contribute to sex differences in language as well.
IX. SEXUAL DIFFERENTIATION OF THE HYPOTHALAMUS & AMYGDALA
As is well known, sexual differentiation is strongly influenced by the presence or absence of gonadal androgen hormones during certain critical periods of prenatal development in many species including humans. However, not only are the external genitalia and other physical features sexually differentiated but certain regions of the brain have also been found to be sexually dimorphic and differentially sensitive to steroids, particularly the amygdala and the preoptic area and medial nucleus of the hypothalamus. Specifically, the presence or absence of the male hormone, testosterone, during this critical neonatal period, directly effects and determines the pattern of interconnections between the amygdala and hypothalamus, between axons and dendrites in these nuclei, and thus the organization of specific neural circuits. In the absence of testosterone, the female pattern of neuronal development occurs.
That various limbic regions, such as the preoptic and medial (ventromedial) hypothalamus are sexually differentiated is not surprising in that it has long been known that this area is extremely important in controlling the basal output of gonadotrophins in females prior to ovulation and is heavily involved in mediating cyclic changes in hormone levels (e.g. estrogen, progesterone). Chemical and electrical stimulation of these nuclei also triggers sexual behavior and even sexual posturing in females and males.
Moreover, In primates, electrical stimulation of the preoptic area increases sexual behavior in males, and significantly increases the frequency of erections, copulations and ejaculations, we well as pelvic thrusting followed by an explosive discharge of semen even in the absence of a mate. Conversely, lesions to these nuclei eliminates male sexual behavior and results in gonadal atrophy.
Similarly, electrical stimulation of the amygdala, the medial division in particular, results in sex related behavior and activity. In females this includes ovulation, uterine contractions and lactogenetic responses, and in males penile erections.
Conversely, damage to the amygdala bilaterally, often results in heightened and indiscriminate sexual activity. For example, primates and other animals (while in captivity) will engage in excessive masturbation and genital manipulation and will repeatedly attempt to copulate even with species other than their own (e.g. a cat with a dog, a dog with a turtle, etc.) regardless of their sex. Hence, with bilateral destruction, animals are not only overly active sexually, but are unable to identify appropriate partners. Conversely, with abnormal activity involving the amygdala, such as due to temporal lobe epilepsy, sensations of sexual excitement, and even sexual behavior sometimes leading to orgasm, may also occur as a function of seizures originating in the temporal lobe.
A. SEX DIFFERENCES IN LANGUAGE & COGNITION
Hence, it thus appears that the limbic system is not only involved in all aspects of emotion, including sexual behavior, and the production of emotional speech, but that these same limbic nuclei may be responsible for sex differences in thought, feeling and even language. For example, it has been argued by that sex differences in language, emotion, and cognitive capability may represent the differential effects of early hormonal influences on various limbic system nuclei as well as within the neocortex. Indeed, the administration of testosterone to infant females during these early critical periods, or the castration of infant males will completely reverse sex differences in behavior and cognition.
For example, it is well known that men, boys, and even male rats demonstrate superior spatial-perceptual abilities, such as in maze learning, as compared to females. If testosterone is not present during these early critical periods, these superiorities are reversed. On the other hand, women and young girls are clearly superior in regard to various aspects of language, including those related to social and emotional functioning. It is likely that these superiorities, like those of the male, are also related to early hormonal influences on limbic organization.
Consider for example intonation and pitch. Women tend to employ 5-6 different variations and to utilize the higher registers when conversing. They are also more likely to employ glissando or sliding effects between stressed syllables, and they tend to talk faster as well. Men tend to be more monotone, employing 2-3 variations on average, most of which hovers around the lower registers. Even when trying to emphasize a point males are less likely to employ melodic extremes but instead tend to speak louder.
B. SEX DIFFERENCES IN EMOTION
As has been demonstrated in a number of recent studies, women are also more emotionally expressive, and are more perceptive in regard to comprehending emotional verbal nuances. This superior female sensitivity even includes the comprehension of emotional faces, and the ability to feel and express empathy. In fact, from childhood to adulthood women appear to be much more emotionally expressive than males in general.
Indeed, given woman's role in rearing children, and the role of the limbic system in promoting maternal care and communication, it seems rather natural that they are much more sensitive to and expressive of these nuances. Presumably, these differences reflect sex related differences in the structure and function of the male vs female limbic system.
Indeed, although sex differences in the structure of the cingulate have not yet been reported, consider for example, the anterior commissure, a bundle of fibers which acts to interconnect the two amygdalas and inferior temporal lobes. This fiber pathway is 18% larger in the female vs the male brain. Given the preimmanent role of the amygdala in emotionality and sound production, as well as evidence indicating that this nuclei is sexually dimorphic, this latter finding of an enlargement in the anterior commissure may be yet another reason why females are more emotionally expressive, receptive, and tend to employ a wider range of melodic pitch when they speak. Moreover, given the intimate role of the amygdala with the hippocampus, it is possible that sexual differentiation of this and other limbic nuclei may be responsible for sex differences in spatial-perceptual abilities and other cognitive differences as well.
X. HIPPOCAMPUS
A. AROUSAL, ATTENTION & INHIBITION
The hippocampus is an elongated structure located within the inferior medial wall of the temporal lobe, posterior to the amygdala and is shaped somewhat like a telephone receiver. It consists of an anterior and posterior region, and is richly interconnected with the septal nuclei (which in some ways acts as a relay nucleus for the hippocampus), as well as the cingulate gyrus and amygdala. And among animals it has been also been found to be sexually differentiated.
Various authors have assigned the hippocampus a major role in information processing, including memory, new learning, spatial mapping of the environment, voluntary movement toward a goal, as well as in attention and behavioral arousal. For example, hippocampal cells greatly alter their activity in response to certain spatial correlates, particularly as an animal moves about in its environment. It is also intimately involved in the encoding and memory storage of spatial, as well as verbal, emotional, and other forms of information. However, few studies have implicated the hippocampus in emotional functioning per se, although responses such as "anxiety" or "bewilderment" have been observed when directly electrically stimulated.
Over the course of evolution the hippocampus has become modified and many of its functions have come to be hierarchically mediated, controlled, or at least, influenced by activity occurring within the neocortex, with which it maintains rich interconnections. Due to this interrelationship the hippocampus is able to monitor as well as exert reciprocal influences over neocortical functioning which it monitors.
For example, when the neocortex becomes highly activated, the hippocampus functions at a much lower level of arousal in order not to become overwhelmed. When the neocortex is not highly aroused, the hippocampus presumably compensates by increasing its own level of arousal so as to tune in to information that is being processed at a low level of neocortical intensity. However, in situations where both the neocortex and the hippocampus become highly aroused and activated, the individual becomes easily distracted, hyper responsive, and overwhelmed, confused, and disoriented. Attention, learning, and memory functioning are also decreased due to this interference in the ability to selectively maintain attention. Situations such as this sometimes sometimes occur when individuals are highly anxious or upset.
Unfortunately, under conditions of extreme and repetitive stress and arousal, the hippocampus may be injured and the victim may suffer varying degrees of memory loss.
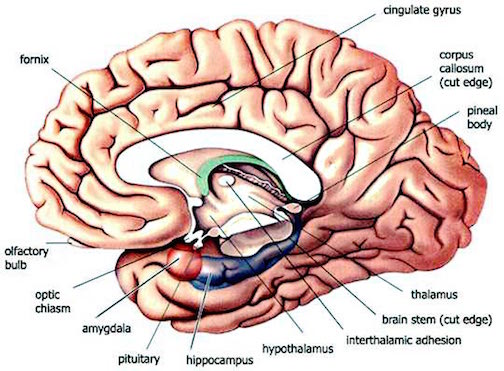
There is also evidence to suggest that the hippocampus may act so as to reduce extremes in neocortical arousal. For example, whereas stimulation of the reticular activating system augments cortical arousal and EEG evoked potentials, hippocampal stimulation reduces or inhibits these potentials such that cortical responsiveness and arousal is dampened.
On the other hand, if neocortical arousal is at a low level, hippocampal stimulation often results in an augmentation of the neocortical evoked EEG potential, thus increasing arousal levels. It is presumably in this manner that the hippocampus can exert influence on what is being processed in the neocortex so as to control selective attention and maintain concentration. Again, this aids in learning and the retention of significant information via selective attention or the filtering of irrelevant forms of input that might otherwise become processed and attended to.
The hippocampus thus prevents the neocortex from becoming overwhelmed or inattentive, and may act to increase neocortical arousal so that it is sufficiently activated. This is because very high or very low states of excitation are incompatible with alertness and selective attention as well as the ability to learn and retain information. When the hippocampus is damaged or destroyed, animals have great difficulty inhibiting behavioral responsiveness or shifting attention. The ability to shift from one set of perceptions to another, or to change behavioral patterns is disrupted and the organism becomes overwhelmed by a particular mode of input. Learning, memory, as well as attention, are greatly compromised. Again, the hippocampus (as well as the amygdala) may be injured under these conditions--a function of the massive release of stress hormones which attack and injure pyramidal cells.
B. LEARNING & MEMORY
The hippocampus is thus associated with learning and memory encoding (e.g. long term storage and retrieval of newly learned information), particularly the anterior regions. Of course, many other brain areas such as the mammillary bodies, dorsal medial nucleus of the thalamus, etc., are also important in memory functioning. Nevertheless, the hippocampus, in conjunction with the amygdala, appears to be preeminent in this regard.
It is now well known that bilateral destruction of the anterior hippocampus results in striking and profound disturbances involving memory and new learning i.e. anterograde amnesia. For example, one such individual who underwent bilateral destruction of this nuclei (H.M.), was subsequently found to have almost completely lost the ability to recall anything experienced after surgery. If you introduced yourself to him, left the room, and then returned a few minutes later he would have no recall of having met or spoken to you. Dr. Brenda Milner has worked with H.M. for almost 20 years and yet she is an utter stranger to him. However, events that occurred for up to two years before his surgery was also somewhat disrupted.
Nevertheless, H.M. is in fact so amnesic for everything that has occurred since his surgery, that every time he rediscovers that his favorite uncle died (years after his surgery) he suffers the same grief as if he had just been informed for the first time. Even so, although without memory for new (non-motor) information, H.M. has adequate intelligence, is painfully aware of his deficit and constantly apologizes for his problem. "Right now, I'm wondering" he once said, "Have I done or said anything amiss?" You see, at this moment everything looks clear to me, but what happened just before? That's what worries me. It's like waking from a dream. I just don't remember...Every day is alone in itself, whatever enjoyment I've had, and whatever sorrow I've had...I just don't remember."
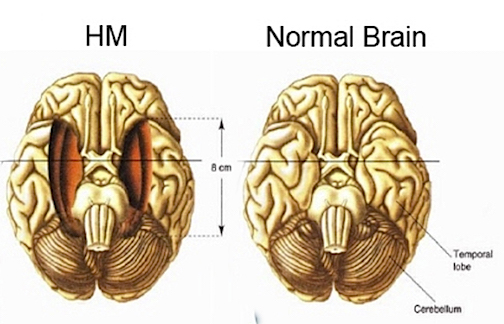
As noted above, presumably the hippocampus acts to protect memory and the encoding of new information during the storage and consolidation phase via the gating of afferent streams of information and the filtering/exclusion (or dampening) of irrelevant and interfering stimuli. When the hippocampus is damaged there results input overload, the brain is overwhelmed by irrelevant stimuli, and the consolidation phase of memory is disrupted such that relevant information is not properly stored or even attended to. Consequently, the ability to form associations (e.g. between stimulus and response) or to alter preexisting schemas (such as occurs during learning) is attenuated.
C. HIPPOCAMPAL & AMYGDALOID INTERACTIONS: MEMORY
The amygdaloid nucleus via its rich interconnections with other brain regions is able to sample and influence activity occurring in other parts of the cerebrum and add emotional color to ones perceptions. As such it is highly involved in the assimilation and association of divergent emotional, motivational, somesthetic, visceral, auditory, visual, motor, olfactory and gustatory stimuli. Thus it is very concerned with learning, memory, and attention, and can generate reinforcement for certain behaviors.
Moreover, via reward or punishment it can promote the encoding, storage and later retrieval of particular types of information. That is, learning often involves reward and it is via the amygdala (in concert with other nuclei) that emotional consequences can be attributed to certain events, actions, or experiences, as well as extracted from the world of possibility so that it can be attended to and remembered. Indeed, the amygdala, in conjunction with the hippocampus, is extremely important in learning and memory, and both are richly interconnected.

The amygdala thus seems to reinforce and maintain hippocampal activity via the identification of motivationally and emotionally significant information and the generation of pleasurable rewards (through action on the lateral hypothalamus). This is because reward increases the probability of attention being paid to a particular stimulus or consequence as a function of its association with reinforcement. As such, events which are positively, or negatively reinforced, are more likely to be remembered; the exception being that high levels of arousal can interfere with memory.
Hence, the hippocampus acts to reduce or enhance extremes in arousal associated with information reception and storage in memory, whereas the amygdala acts to identify the social-emotional-motivational characteristics of the stimuli as well as to generate (in conjunction with the hippocampus) appropriate emotional rewards so that learning and memory will be reinforced. Thus, we find that when both the amygdala and hippocampus are damaged, or under conditions of prolonged and repetitive stress, striking and profound disturbances in memory functioning result.
D. VISUAL & VERBAL MEMORY
It is now very well known that lesions involving the inferior temporal lobes and the amygdala/hippocampus of the left cerebral hemisphere typically produce significant disturbances involving verbal memory. Left sided damage disrupts the ability to recall simple sentences, complex verbal narrative passages, or to learn verbal paired-associates or a series of digits.
In contrast, right temporal, amygdala-hippocampal destruction typically produces deficits involving visual and spatial memory, such as the learning and recall of geometric patterns, visual mazes, human faces, or even where some object was placed the night before. Right sided damage also disrupts the ability to recall olfactory stimuli, emotional sounds and passages, or sounds from the environment.
Hence, the left amygdala/hippocampus is highly involved in processing and/or attending to verbal information, whereas the right amygdala/hippocampus is more involved in the learning, memory and recollection of non-verbal, visual-spatial, environmental, emotional, motivational, and facial information. However, as noted above, the limbic system, including the hippocampus, is sexually differentiated, which in turn appears to effect the ability to attend to and recall spatial vs emotional and verbal information. In this regard, the male limbic system appears to have conferred an advantage in the processing of spatial information, whereas the female limbic system is more adept at expressing, processing and possibly recalling emotionally laden visual and verbal stimuli.






