





Rhawn Joseph, Ph.D.
BrainMind.com
THE NEOCORTEX
Perception, cognition, fine-motor expression, and computational processing, is made possible by neurons, the majority of which are pyramidal neurons. Most pyramidal neurons are located in the neocortical mantle of the lobes of the brain, which gives this outer coating its grayish appearance. Over 90% of the gray matter is located in the neocortex. The neocortex ("new cortex") likely first began to evolve between 300 million to 450 million years ago.
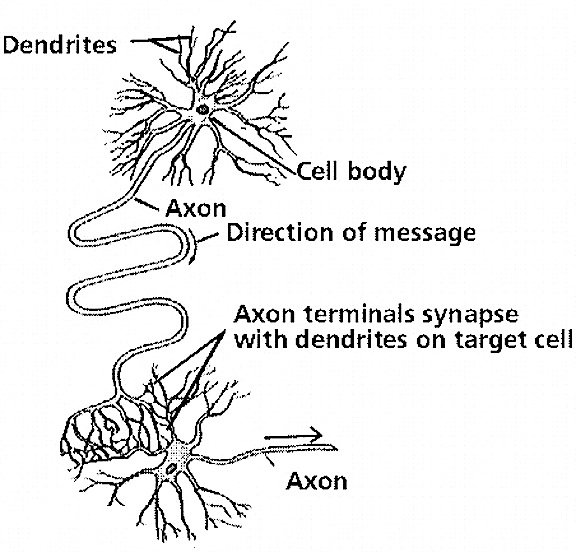
Those neurons which project to neurons in the next column or to those neurons in an upper or lower layer, are referred to as local circuit neurons. Those which project from the neocortex to the brainstem, or from one half of the brain to the other, are referred to as long distance neurons (Peters & Jones, 2010).
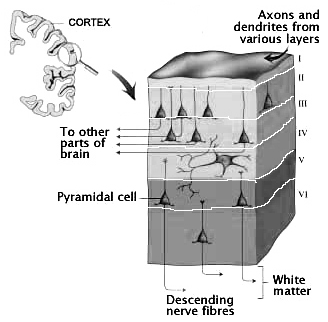
Classically, the neocortex is said to consist of six to seven layers when in fact it consists of numerous layers which vary depending on brain area (Braak & Braak, 2017; Peters & Jones, 2010; Ramon y Cajal, 1902/1955; Rose, 1926). For example, the deepest layer, neocortical layer VI, consists of two distinct layers (VIa and VIb). In the occipital lobe, three additional layers (i.e. sublayers) can be distinguished within layer IV (which also receives considerable thalamic input and is very thick). By contrast, within the motor areas of the frontal lobe, layer IV is exceedingly thin (as there is comparatively minimal thalamic input), whereas layer V is exceedingly thick, It is layer V of the frontal motor areas which contribute the bulk of axons that form the descending corticbulbar, corticopontine, and corticorubral brainstem pathways which establish contact with cranial nerve and sensory and spinal motor neurons (Brodal, 2011; Catsman-Berrevoets, 2017). Likewise in the temporal neocortex layer V is relatively thick as are layers I and VI (since much of the temporal lobe is association and assimilation cortex). As noted, the entorhinal cortex, the "gate way to the hippocampus" and which is located along the medial surface of the temporal lobe, consists of between 7 and 8 layers (Braak & Braak, 2017; Ramon y Cajal, 1902/1955; Rose, 1926).
Hence, the thickness, layering, and composition of the human neocortex varies from lobe to lobe and actually consists of from 7 to 9 (or more) layers rather than 6. For our purposes (and throughout this book) we will described the neocortex as having 7 layers.


Layer III is the Pyramidal Layer and consists of medium pyramidal cells which project axons to distant areas of the neocortex. Hence, the neurons of layer III can be considered long distance neurons.
Layer IV, the Internal Granular Layer, has a granular appearance and consists of small pyramidal, granule, and stellate (starshaped) cells and receives massive axonal projections from the thalamus. These neurons are predominantly local circuit, and project to adjacent columns and layers. That is, upon receiving and analyzing thalamic input, the neurons of layer IV transfer this data to adjacent neurons for additional analysis. Because the primary, secondary and association sensory areas receive considerable thalamic input, layer IV is relatively thick--except in the motor cortex.
Layer V is the Ganglionic Layer and consists of large and medium size pyramidal cells, including, in primary motor cortex (Brodmann's area 4) the giant cells of Betz. The pyramidal neurons of layer V are long distance neurons, and give rise to descending axons which form the corticospinal, pyramidal, corticobulbar, corticopontine, and corticorubral brainstem pathways which establish contact with cranial nerve and sensory and spinal motor neurons (Brodal, 2014; Catsman-Berrevoets, 2010). It is these "pyramidal" and cortico-spinal neurons which make purposeful, fine motor movement possible. Approximately 31% of the corticospinal tract arises from the pyramidal cells located in the primary motor areas 4, with the remainder arising from the frontal motor associations areas 6, 8, and the primary somesthetic areas 3,1,2, with a scattering of fibers being contributed by the occipital and temporal lobe, as well as limbic system structures.


Intra-cortical Neurons
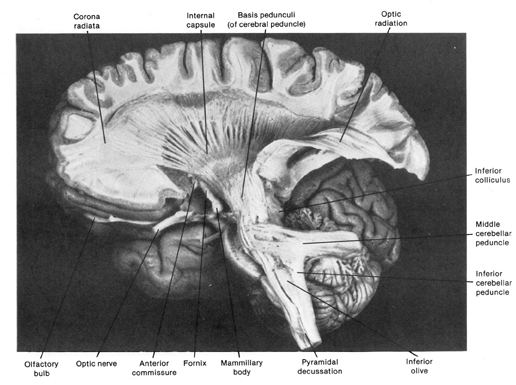
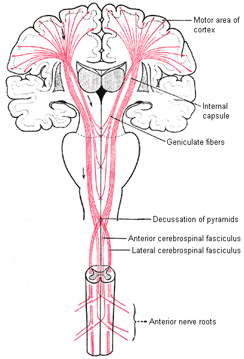
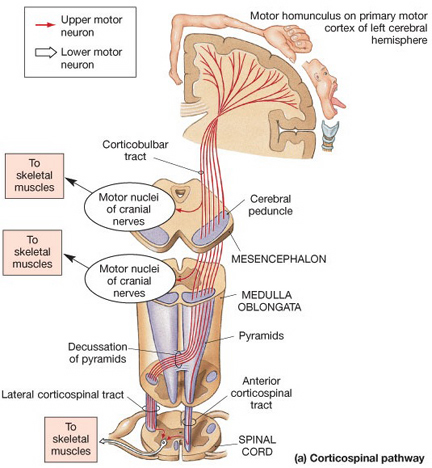
Layer VIa is the Multiform Layer and contains pyramidal, fusiform, and spindle shaped cells, whereas Layer VIb consists of predominantly of spindle shaped cells. These are predominantly local circuit neurons, and receive considerable input from the brainstem.
The Cytoarchitextural Neuronal, & Chemical Organization of the Neocortex
Korbinian Brodmann detailed the regional variation in the cytoarchitectural organization of the cortex, and conducted detailed comparative studies of numerous species, each of which displays common as well as varying patterns of cytoarchitexture and gyral folding. Based on these cytoarchitextural differences and commonalities, Brodmann divided the cortex into distinct regions and created cytoarchitextural maps of the brains of a variety of species, including humans. For examples, Brodmann's area 17 is synonymous with the primary visual cortex, whereas Brodmann's areas 3,1,2, denote and identify the primary somesthetic receiving areas.
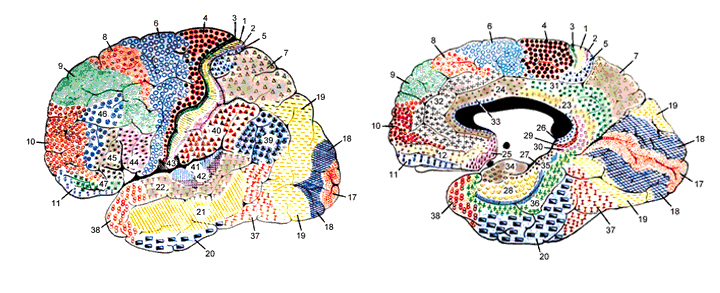
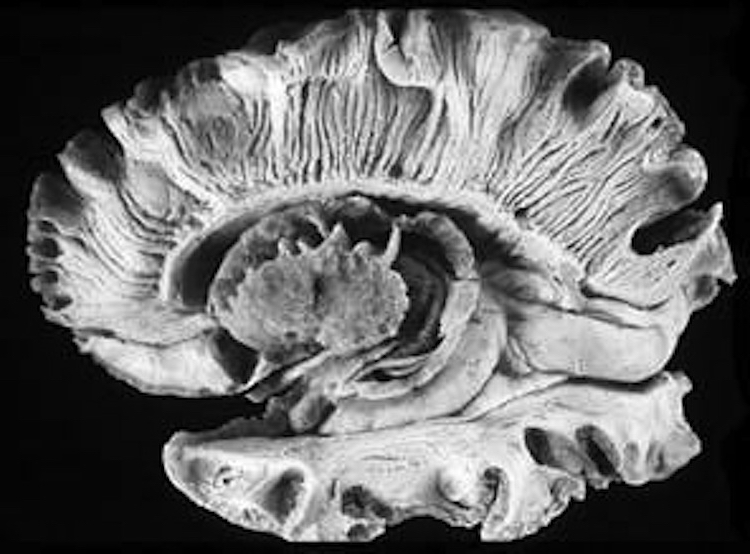
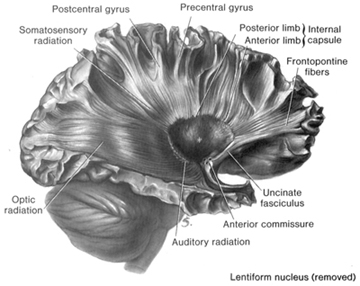
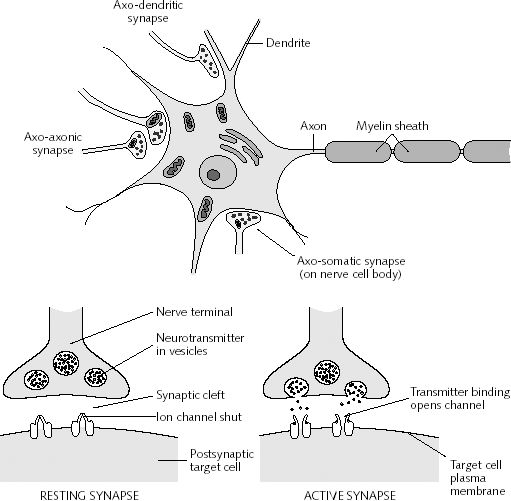
However, although these area differ in regard to organization, what they share in common is a preponderance of pyramidal cells. As noted, pyramidal cells are also the largest and are more numerous than any other neocortical neuron (Peters & Jones, 2010). Pyramidal neurons account for up to 3/4 of all neocortical cells. Pyramidal neurons also serve as both local-circuit and long-distance neurons and generally receive two types of synaptic contacts referred to as Gray types I and II which differ in synaptic morphology and (respectively) excitatory vs inhibitory influences (Peters & Jones, 2010). However, almost all pyramidal cell are excitatory and use glutamate and aspartic acid as transmitters (Tsmoto, 1990).
Pyramidal cells can also be classified as Golgi I and II cells. However, of all neocortical neurons, only 10% are Golgi type I neurons, the main source of long-distance (excitatory) axons, the majority are interneurons, i.e. local circuit neurons which in turn provide almost 90% of cortical axons and dendrites. Approximately 95% of Golgi type I long distance axons interconnect distant regions within the same hemisphere and only about 5% cross the corpus callosum, the fiber pathways which link the right and left hemisphere (Peters & Jones, 2010).
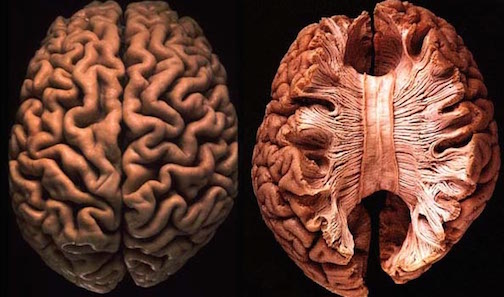

In addition to glutamate and GABA, neocortical neurons contain and respond to peptides, including substance P, corticotropin releasing factor, and opiates. The peptide containing neurons tend to congregate in layers II, III, and IV (Jones & Hendry, 1986).
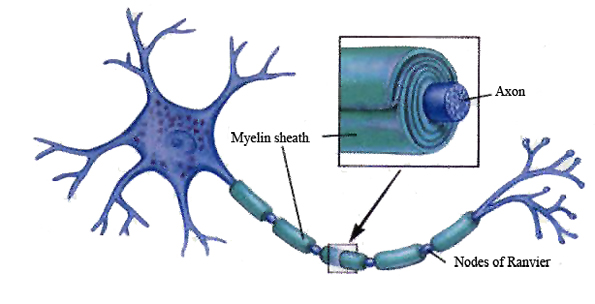

Glia and non-neuroglia elements make up almost 70% of the volume of the neocortex. Of the remainder, 22% consists of axons and dendrites, with the body (soma) of the neuron comprising only 8% (Peters & Jones, 2010).
REFERENCES










