





BrainMind.com
Functional Neuroanatomy of the Parietal Lobe Primary Sensory Receiving Area
Primary Somesthetic Receiving Areas
The parietal lobes are not a homologous tissue but consists of cells which are responsive to a variety of divergent stimuli, including movement, hand position, objects within grasping distance, audition, eye movement, pain, heat, cold, as well as complex and motivationally significant visual stimuli (Aoki et al., 2018; Cohen et al. 2012; Deibert et al., 2009; Dong et al. 2014; Lam, et al., 2018; Lebedev et al. 2011; Lin & Sessle 2017; Pred'Homme & Kalaska 2014; Remy et al., 2009; Snyder et al., 1998).
For example, area 7 of the superior parietal lobe receives considerable visual input, particularly from the lower visual fields and periphery--information which it receives not only from the visual cortex but from extra-retinal pathway. Area 7 also receives complex somesthetic stimuli regarding the hand and objects beyond and within reaching distance. Moreover, different neurons in different regions of area 7 perform somewhat different functions. Even the subareas within the primary somesthetic neocortex, areas 3ab,1, 2, respond to different stimuli and receive projections from different subregions within the thalamus, which in turn receive different forms of input from skin, joints and muscles.
Because it serves so many diverse yet related functions, damage to the parietal lobe can therefore result in a variety of disturbances. This includes abnormalities involving somesthetic and pain sensation, the body image, visual-spatial relations, temporal-sequentual motor activity, language, grammar, numerical calculation, emotion, and attention, depending on which area has been lesioned as well as the laterality of the damage.
PARIETAL TOPOGRAPHY
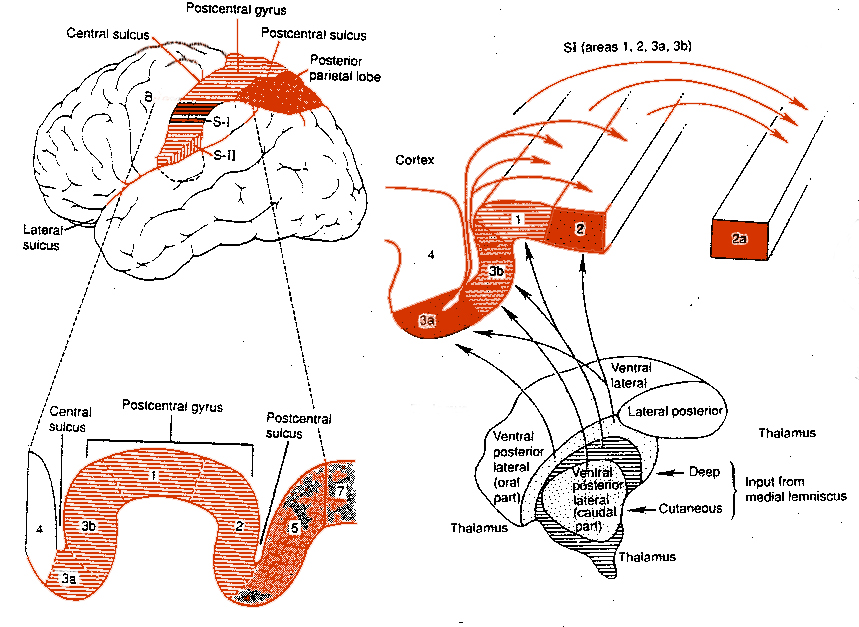
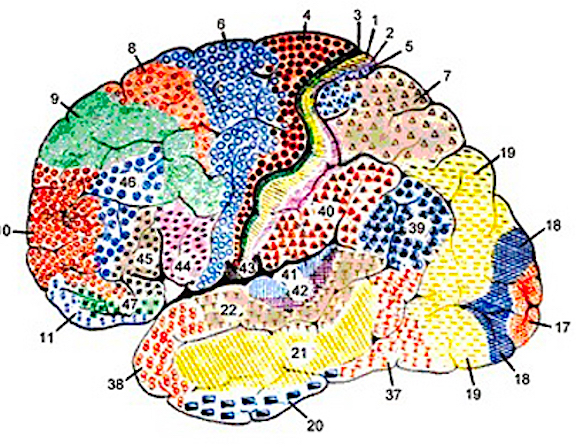
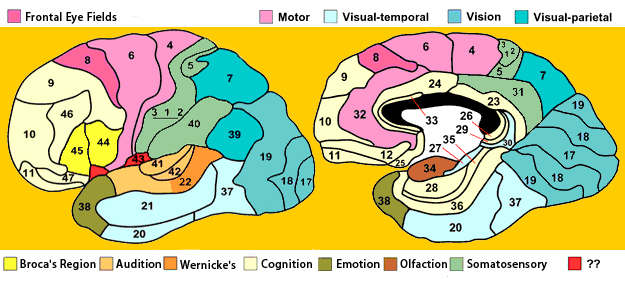
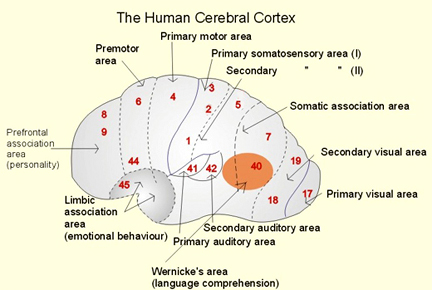
There are nine major somesthetic areas within the parietal lobe, such that the primary, association, and assimilation areas actually consist of numerous subareas. Broadly, and most generally, however, the parietal lobe may be subdivided into a primary receiving area (involving Brodmann's areas 3ab,1,2) within the post central gyrus, an immediately adjacent somesthetic association area (Brodmann's area 5ab), a polymodal (visual, motor, somesthetic) receiving area located in the superior-posterior parietal lobule (area 7ab), a granular insular area which is located in the inferior convexity and encompasses part of the marginal gyrus, and a multimodal-assimilation area within the inferior parietal lobule (areas 7, 39, 40) which encompasses the angular and supramarginal gyrus.
THE PRIMARY SOMESTHETIC RECEIVING AREAS
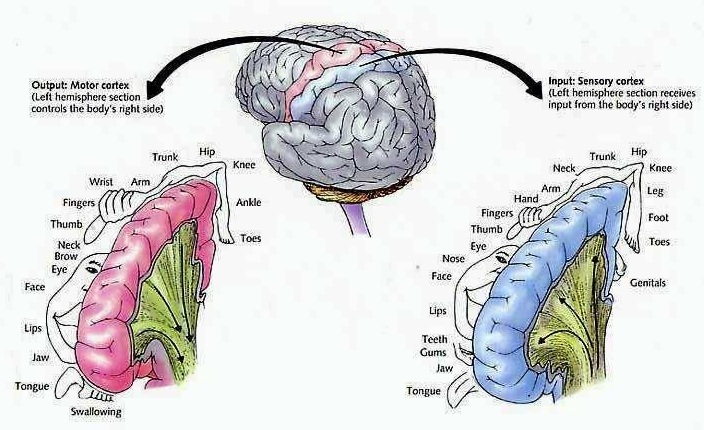
The primary somesthetic areas consists of three narrow strips of tissue (areas 3ab, 1, 2) which differ histologically, in architectural composition, and in sensory input. Moreover, each of these areas maintains a complete and independent representation of the body (Kaas, 1993).
Specifically, area 3a receives input from the muscles spindles (group IA muscle afferents) and can also signal muscle length (e.g. flexion or extension) whereas areas 3b receives cutaneous stimuli. Hence, area 3 appears to maintain cutaneous and muscular maps of the body. However, almost all of the cells in area 3ab receive input only from the contralateral half of the body. Hence, only half the body is represented.
These two maps, however, are also semi-independent, and in some respects they do not directly correspond to the location of body parts along the body surface, but instead are organized in regard to those parts which most frequently interact. That is, not only are certain body parts more greatly represented in accordance with their sensory importance, but in area 3b, for example, the hands, fingers, and jaw and mouth are juxtaposed--a function of the interaction of the hand and mouth when eating food (Kaas, 2013).
Information received and processed in area 3 is relayed to the immediately adjacent areas 1, and 2; each of which also contain a specialized spatial map of the body (Kaas, 1993; Kaas et al. 1981; Lin et al. 2014; Sur et al. 1982). For example, area 1 appears to maintain an overlapping cutaneous-joint body map (Evarts, 1969; Mountcastle & Powell, 1959; Schwartz et al. 1973). Area 2 maintains a map of the joint receptors and can signal the position and posture of the limbs based on input from the muscle spindles. Hence, the somesthetic cortex maintains four independent maps of the body.
Moreover, within this tiny expanse of tissue there is a sequential hierarchical convergence of input from areas 3, to 1, onto area 2. That is, information is analyzed and then passed from area 3 to 1, and from 3 and 1 onto area 2, as well as from area 3 to area 2. Therefore, a single neuron in area 2 receives multiple input from several cells in area 1 and 3, as well as vestibular input.
This organization is evident anatomically and as based on single cell recording and evoked potentials. For example, evoked potentials appear in area 1 about 5 msec, after they appear in area 3b. However, the degree of activation is also dependent on the attentional state and degree of arousal. With minimal attention to the source of input, there is minimal activation, which is why sensations from the clothing, shoes, or while sitting, lying down, and so on, can rapidly fade from consciousness.
Together these four strips of tissue comprise an interactional functional unit and are responsive to touch, texture, shape, motion, and the direction of stimulus movement, including temporal-sequential patterning, and can directly monitor the position and movement of the extremities (Cohen et al. 2014; Lebedev et al. 2014; Levitt & Levitt, 2008; Lin et al. 2014; Pred'Homme & Kalaska 2014; Mountcastle, 1957; Warren et al. 2016ab; Whitsel et al. 2012). Many cells are also responsive to changes in temperature as well as the presence of noxious stimuli applied to the skin.
Because the majority of these neurons receive input concerning pressure, light touch, vibration, the movement of joints, and muscular activity (Cohen et al. 2014; Lebedev et al. 2014; Levitt & Levitt, 2008; Prud'Homme & Kalaska 2014; Mountcastle, 1957) they can signal and determine whatever posture or position the body is in as well as the amount of force or pressure being exerted by the limbs (Jennings et al. 1983), i.e. if carrying or lifting some object. Conversely, via the reception and analysis of this input an individual can detect an insect crawling up or down their leg, the direction it is moving, as well as determine the position of their arms and legs without looking at them.
Nevertheless, predominantly elementary and simple contralateral somesthetic information is processed in this region (Lin et al. 2014; Prud'Homme & Kalaska 2014). Electrical stimulation of the primary somesthetic area gives rise to simple, albeit well localized sensations on the opposite half of the body (Penfield & Boldrey, 1937; Penfield & Jasper, 1954; Penfield & Rasmuseen, 1950) such as numbness, pressure, tingling, itching, tickling and warmth.
BODY IMAGE REPRESENTATION
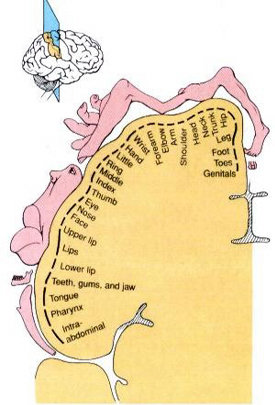
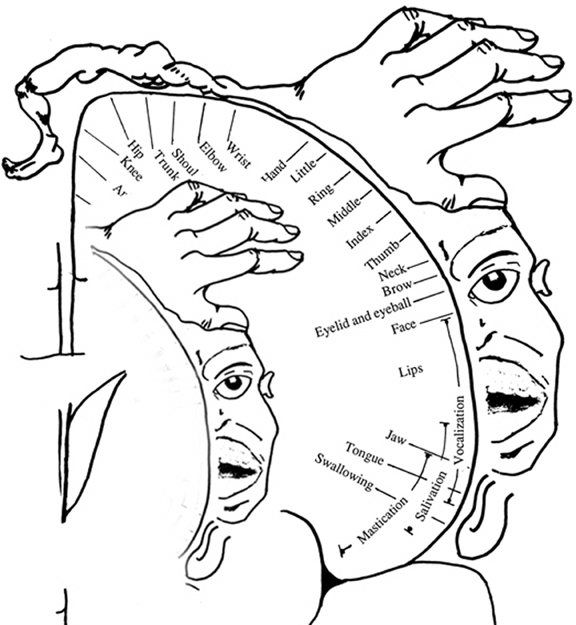
The primary receiving areas for somesthesis continues up and over the top of the hemisphere and along the medial wall where the lower half of the body is represented. Specifically the rectum, genitals, foot and calf are located along the medial wall, the leg along the superior surface of the hemisphere, and the shoulder, arm, hand and then face along the lateral convexity (Penfield & Boldrey, 1937; Penfield & Jasper, 1954; Penfield & Rasmussen, 1950).
Body parts are also represented in terms of their sensory importance, i.e. how richly the skin is innervated. For example, more cortical space is devoted to the representation of the mouth, fingers and the hand than to the elbow or trunk (Warren et al. 2016). In fact, the area devoted to representation of the fingers is 100 times larger than the area devoted to the trunk. Because of this the cortical body map is very disorted. However, as noted, some areas are also juxtaposed, such as the hand and mouth area.
In summary, the primary receiving area receives very precise information regarding events occuring anywhere along the internal/external body and responds to converging inputs from muscle spindles, cutaneous and joint receptors, as well as proproceptive and vestibular stimuli. In this manner, not only the body but the global properties of objects held in the hand can be determined (Iwamura & Tanaka, 1978); i.e. stereognosis.
FUNCTIONAL LATERALITY
As detailed in Chapter 10, there is clear evidence that the right parietal area is dominant in regard to many aspects of somesthetic information processing. Hence, neurons in this half of the brain appear to be more sensitive and more responsive and to more greatly monitor events occuring on either half of the body, but particularly the left. In fact, this relationship was noted over 150 years ago by Weber.
According to Weber (1834/1977), the left half of the body exceeds the right in regard to most forms of tactual sensitivity. The left hand and the soles of the left foot, as well as the left shoulder are more accurate in judging weight, have a more delicate sense of touch and temperature, such that "a greater sense of cold or of heat is aroused in the left hand" (p. 322). That is, the left hand judges warm substances to be hotter, and cold material to be colder as compared to the right hand, even when both hands are simultaneously stimulated.
SOMESTHETIC AGNOSIAS
Surgical or other forms of destruction involving the primary somesthetic receiving areas results in a complete, albeit, temporary loss of sensation from the entire half of the body (Russell, 1945). Longer term effects include elevation of sensory detection thresholds, loss of position and pressure sense, including two-point discrimination, and a greatly reduced ability to detect movement of the fingers. In addition, the capacity to determine texture, shape, temporal-sequential patterning, or to recognize objects by touch or to discriminate among different forms or their properties, e.g. size, texture, length, shape and stereognosis is significantly attenuated (Corkin et al. 1970; Curtis et al. 2012; LaMotte & Mountcastle, 1979; Randolf & Semmes, 2014). Passive, (non-movement) sensation is less impaired. In some instances, over time a remarkable recovery of somesthetic discrimination sense may be observed (Semmes, 1973).
Nevertheless, even with complete removal of the post-central gyrus, stimuli applied to the face are much better perceived as compared to the same stimuli applied to the hand. Conversely, lesions which spare the hand area of the post-central gyrus, but which destroy the remaining tissue, will result in mild or no permanent sensory deficits when the hands are tested (with the exception of stereognosis; Semmes, 1965). However, sensation across other body parts will be impaired. Hence, when testing for parietal lobe dysfunction, not only the face and hands, but other body parts should be examined.
As noted, this region is also highly concerned with motor functioning as it is extensively interconnected with the primary motor area and gives rise to almost 1/3 of the cortico-spinal tract. Hence, damage to this region can give rise to motor disturbances such as paresis accompanied by hypotnonia, and/or produce inaccuracy and reduced speed of movement (Cole & Glees, 2014; Luria, 1980). The ability (or will) to initiate movement may also be reduced.
REFERENCES










