





Rhawn Joseph, Ph.D.
BrainMind.com
Simple and complex visual and central/foveal analysis is one of the main functions associated with the occipital lobe (Kaas & Krubitzer 1991; Sereno et al. 2016; Zeki, 2007). However, like the frontal, temporal, and parietal lobes which respond to and process information from a number of modalities, the occipital lobe contains neurons which respond to vestibular, acoustic, visual, visceral, and somesthetic input (Beckers & Zeki 2014; Ferster, et al., 1996; Horen et al., 1972; Jung, 1961; Morrell, 1967; Pigarev 2014; Sereno et al. 2016; Zeki. 2007).
THE PRIMARY VISUAL CORTEX
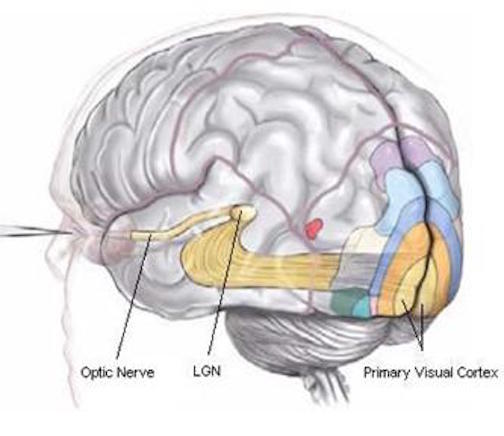
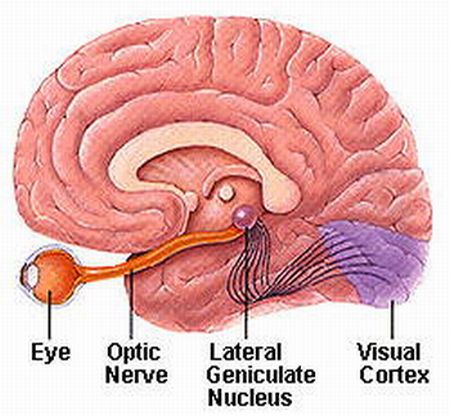
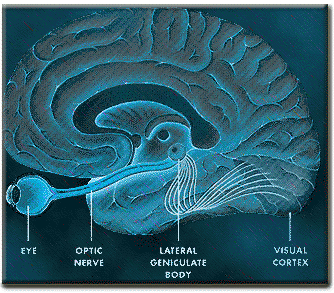
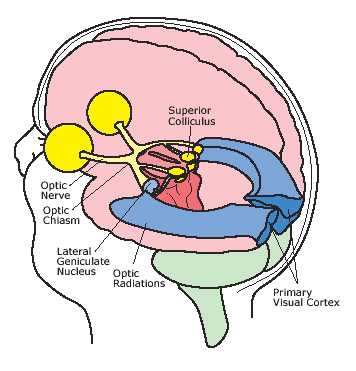
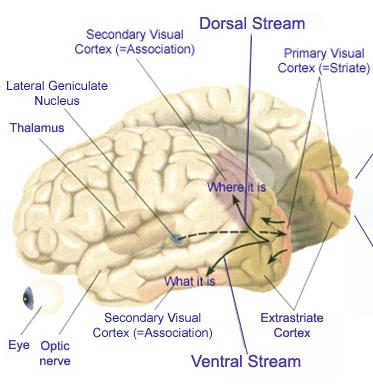
In primates and those carnivores with their forward facing eyes, axons from the retina form the optic nerve, are organized so that visual information from the same points in visual space can be combined and rerouted at the optic chiasm, and then directed so that all information arising from the left vs right half of the visual fields are directed to the right vs left half of the occipital lobes and visual cortex. That is, visual information arising in the nasal portion of the right eye, is combined with that from the lateral-temporal portion of the left eye (and vice versa), such that the primary visual area in the right hemisphere receives information from the left half of visual space. However, this information is first received in the lateral geniculate nucleus of the thalamus, each layer of which receives input from only one eye (Casagrande & Joseph, 1978, 1980), the bulk of which is then relayed to the primary visual cortex.
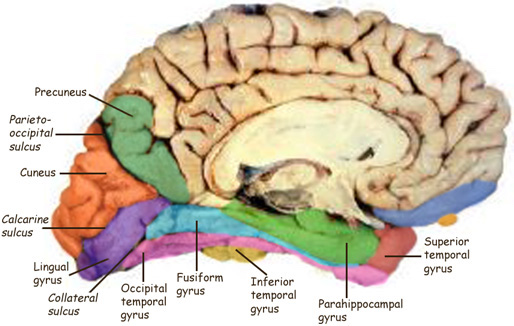
The primary visual receiving area (i.e. striate cortex, area 17) is located predominantely within the medial walls and floor of the calcarine sulcus and extends around the lateral convexity. Area 17 is also characterized by rather thin cortical layers, particularly layers II and II, and by its striped appearance which is due to the structure and composition of layer IV. That is, layer IV is divided into three sublayers, with the middle layer containing a rather thick band of cortex (the band of Baillarger/Gennari) which is visible to the naked eye.
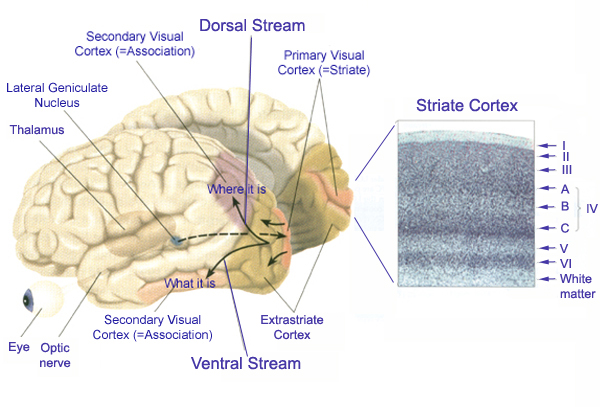
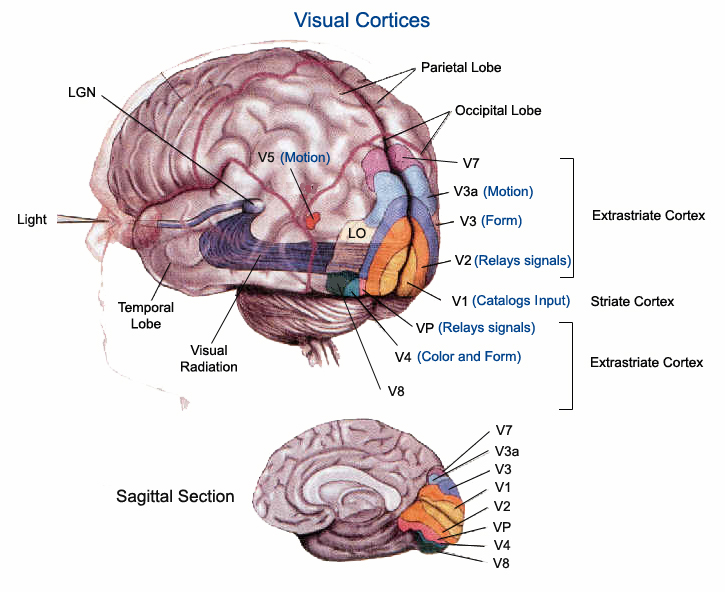
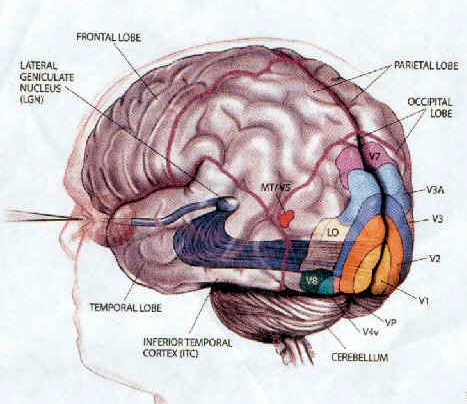
The association cortices (areas 18 & 19) are also located medially and along the lateral convexity. However, there are also extra-striatal visual association and assimilaton areas located in the superior parietal (area 7) and inferior and middle temporal lobes (Kaas & Krubitzer 1991; Nakamura et al. 2014; Sereno et al. 2016; Tovee et al. 2014).
NEOCORTICAL COLUMNAR ORGANIZATION
Throughout the striate cortex neurons with similar receptive properties are stacked in columns (Hubel & Wiesel, 1968, 1974). Indeed, one column of cells may respond to a certain visual orientation and the cells in the next column to an orientation of a slightly different angle. Moreover, columns exist for color (Zeki, 1974), location, movement, etc. In addition, since certain cells respond predominantly to input from one eye, there are ocular (eye) dominance columns as well (Hubel & Wiesel, 1968, 1974). A similar columnar arrangement in regard to somesthetic input is maintained in the parietal lobe.
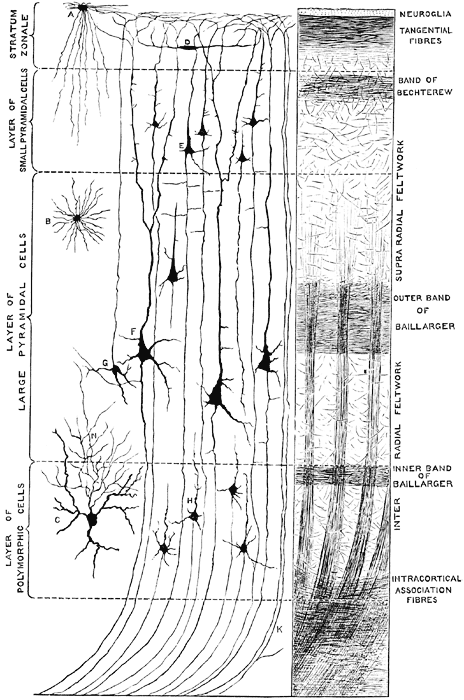
Within these ocular dominance columns, the visual cortex is also organized so that they match and parallel input received within the retina, thus creating a retinotopic map. That is, adjacent cells in these columns receive input from adjacent cells in the retina. However, there is also almost a 50% overlap between columns, such that there are shared receptive fields, such that single cells can perceive more than one point in space--which in turn allows for a smooth transition which making eye and head movements. Indeed, neurons of a particular column, although communicating predominantly with those in the same column, also communicate with immediately adjacent columns, such that a considerable amount of parrallel communication occurs (Dow, 1974; Kaas & Krubitzer 1991). That is, information is anlyazed vertically and horizontally. so as to create a series of superimposed mosaics of the visual word.
SIMPLE, COMPLEX, LOWER- & HIGHER ORDER HYPERCOMPLEX FEATURE DETECTORS
The visual cortex is made up of a variety of cell types each of which is concerned with the analysis of different visual features (Ferster, et al., 1996; Hubel & Wiesel, 1959, 1962, 1968; Kaas & Krubitzer 1991; Sereno et al. 2016). These include simple, complex, and (higher & lower order) hypercomplex cells which are distributed disproportionately throughout areas 17,18, 19.
To briefly summarize, simple cells appear to be involved in the initial analysis of incoming visual cortical input, and are most sensitive to slowly moving stimuli. They are found predominantly within area 17 and in layer IVa,b,c,. Some are sensitive to stimuli moving in one direction, whereas others may respond to stimuli moving in any direction. In fact, almost 95% of the neurons in area 17 are responsive to stimuli moving only in one direction, but not the direction of movement. In addition, simple cells are responsive to the particular position and orientation a stimulus may take. However, for a simple cell to fire, a stimulus must assume a specific orientation and position.
Simple cells relay this processed information to the far more numerous complex cells which are found predominantly in layers II and III and V, which interact and communicate with one another including with layer IV which receives thalamic input. Each complex cell receives input from several simple cells. Complex cells are also concerned with orientation of the stimulus. However, these cells are more flexible and will respond and analyze a stimulus regardless of its particular orientation. These cells via the combined input from simple cells, are probably involved in the earliest stages of actual form perception, i.e. the determination of the outline of an object. A considerable number of complex cells receive converging input from both eyes, the remainder being monocular. Complex cells are found predominantly within area 18.
Hypercomplex cells are concerned with the analysis of discontinuity, angles and corners, as well as movement, position, and orientation. That is, these cells respond selectively to certain visual configurations and thus act so as to determine precise geometric form. It is also via the action of these cells (in conjunction with visual neurons in the temporal lobe) that the first stages of visual closure are initiated. This in part requires that the functional activity of these cells be suppressed such that when presented with an incomplete figure these cells are overridden and the brain is able to "fill in the gaps" in stimuli perceived. It is also for this reason that one does not notice his or her "blind spot"; it is filled in. Hypercomplex cells are found predominantly within area 19.
STRIATE CORTEX: AREA 17
The primary visual cortex, area 17, is located predominantly within the medial walls of the cerebral hemispheres, extending only minimally along the lateral convexity. This area is often referred to as "striate" because the incoming fibers from the optic radiations form a stripe along the cortical surface which can be seen by the naked eye. Areas 18 and 19 do not have this striped appearance.
Like the primary motor and somesthetic cortices, a greater degree of cellular representation is maintained for those areas which are the most densely innervated and of the most sensory importance, i.e., the fovea (Daniel & Whitteridge,2011; Hubel & Wiesel, 1979; Kaas & Krubitzer 1991). Indeed, the central part of the retina has a cortical representation which is 35 times more detailed than that of the periphery. This is particularly important in that the fovea contains cells which are most sensitive to the detection and representation of form.
Although all neuron types are found within the striate cortex, simple and complex cells predominate. Hence, the primary receiving area is predominantly involved in the analysis of color, slow movement, position, and orientation (Zeki, 2007); i.e. the most elementary aspects of form and visual stimulus perception.
The primary visual cortex, however, receives fibers from non-visual brain areas as well. These include brainstem nuclei, the pontine and mesencephalic reticular formation, the lateral amygdala, and lateral hypothalamus (Doty, 1983; Tigges et al. 2013). Processing in the primary region can thus be enhanced or diminished via reticular influences and emotional-motivational concerns. In this manner, if a stimulus is emotionally significant greater visual attention will be directed at the object.
Conversely, bilateral destruction of area 17 can result in loss of visual recognition capabilities -even with sparing of the association areas (Humphrey & Weiskrantz 1967; Weiskrantz & Cowey 1963). However, awareness of moving objects and visual-spatial orientation is preserved. Visual preservation with primary occipital destruction has been referred to as "blind sight" (see below).
HALLUCINATIONS.
Electrical stimulation, tumors, seizures, or trauma involving the striate cortex may produces simple visual hallucinations, such as sparks, tongues of flames, colors and flashes of lights (Penfield, 1954; Tarachow, 1941). Objects may seem to become exceedingly large (macropsia) or small (micropsia), blurred in terms of outline, stretched out in a single dimension, or colors may become modified or even erased (Hecaen & Albert, 1978). Sometimes simple geometric forms may be reported. Usually the hallucination is restricted to one half of the visual field. That is, if the seizure is in the right occipital lobe, the hallucination will appear in the left visual field.
Although elementary hallucinations are usually associated with abnormalities involving the occipital lobe they may occur with temporal lobe lesions or electrical stimulation (Penfield & Rasmussen, 1950; Tarachow, 1941).
REFERENCES










