





Sexual Differentiation Of The Body & Brain (From: Neuropsychiatry, Neuropsychology, Clinical Neuroscience, Academic Press 2000)
Rhawn Gabriel Joseph, Ph.D.
BrainMind.com
SEXUAL DIFFERENTIATION OF THE BODY & BRAIN
There is considerable confusion and misinformation about sexuality. Morality, religion, or politics often trumps biology, and in consequence, ignorance and intolerance reigns.
Sexuality, gender identity, the appearance of the body, and sex-specific behaviors and sex differences in cognition are influenced by genetics, hormones, and the environment. One can be genetically female, yet be born with male-appearing genitalia. One can be genetically male, yet be born "homosexual" or with a "female" brain and body. Its not a matter of choice; its biology. On the other hand, the environment interacts with biology, and gene selection; and thus all manner of variations in sexuality and sexual desire are the result and which may be guided by choice, or vice versa.
The Genetics of Sex
The genetics of sex are determined by the absence or presence of the "male" Y chromosome. No matter how many "female" X chromosomes may be inherited, if there is a single Y chromosome present (e.g. YXXX), genetically the animal or human is "male."
The genetics of gender in turn predisposes the Zygote/Fetus, after the conception to differentiate in either a male of female pattern. However, other factors also play a major role, including maternal stress and the presence or absence of androgens such as testosterone.
Ovaries & Testes
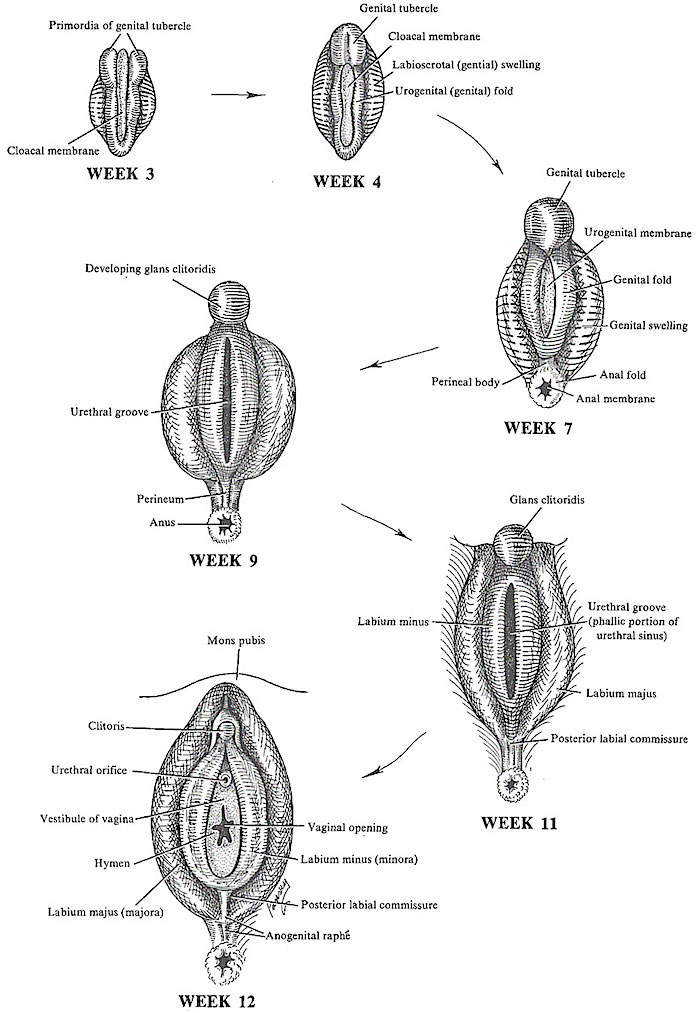
The primordial gonads have the capability of becoming ovaries or testes. Thus the urogenital and the external genitalia are similar in males and females up to the 9th week of fetal development.
However, in the genetically YX fetus, testicular tissues sometimes begin to form in the embryo as early as the 7th week. In the XX fetus the gonads begin to differentiate into ovarian tissue around the 8th week.
However, in part, this differentiation process if dependent on the presence or absence of the "male" hormone, testosterone.
In the XY fetus, testosterone appears to have several sources, including the primordial testes. Yet another source may be the placenta, which is stimulated to produce steroidal substances and gonadotropins at 12 weeks of fetal age. That is, the fetal Y chromosome somehow interacts with and triggers the plancenta to produce gonadotropins which in turn influences early fetal gonadal differentiation and the ability to produce testosterone.
Mullerian & Wolfian Ducts
The genetically male and genetically female fetus posses Mullerian & Wolfian Ducts which in turn will differentiate or degenerate and become the female vs male sex and reproductive organs. The Mullerian ducts give rise to the female reproductive system and the Wolfian ducts give rise to the male.
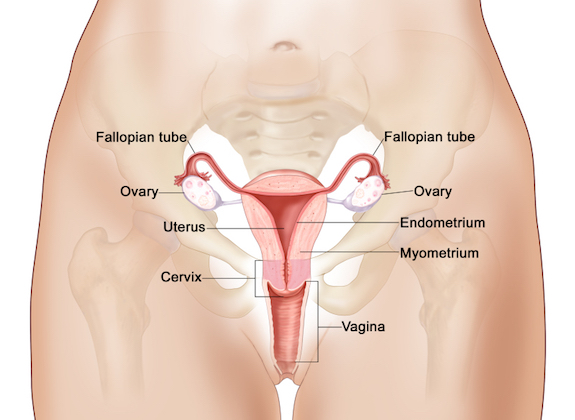
The Mullerian ducts begin to differentiate in the human embryo at around 2 months. The Mullerian ducts differentiate into fallopian tubes, the uterus and the upper part of the vagina. If androgens (male hormones) are present, the Mullerian ducts degenerate.
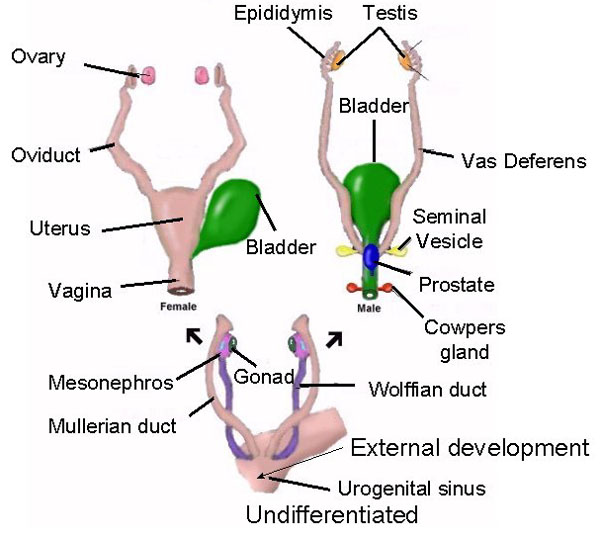
In the male fetus the wolffian duct is dependent of the presence of androgens (testosterone) and becomes the vas deferens system and seminal vessicles which are involved in sperm production and ejaculation. In the female fetus, the wolffian ducts degenerate.
In the male fetus, the wolffian ducts forms the vas deferens and the seminal vesicle. This differentiation is dependent of high local concentration of testosterone, during a "�critical�" period during which the wolffian duct is sensitive, i.e. around 2-3 months of fetal age. In the female fetus, the Wolffian duct degenerates.
The crucial role of testosterone has been demonstrated in a number of species. For example, if the gonads are removed in the XY fetus before differentiation, the wolffian ducts will degenerate and the Mullerian ducts will instead grow and differentiate into fallopian tubes, uterus and the upper part of the vagina (24). If androgens are injected into female fetuses, the mullerian ducts will degenerate and the wolffian ducts will in turn differentiate into the male reproductive system.
However, it is not just the body, but the brain which differentiates into a male vs female pattern. In fact, there is a male, female, and homosexual limbic system. This is important because the limbic system controls all aspects of social and emotional functioning including sexuality and sexual orientation.
MATERNAL STRESS & SEXUAL DIFFERENTIATION
Because of the Mullerian and Wolfian Ducts and the limbic system are sensitive to circulating stereoids, chronic maternal stress may in fact induce steroidal alterations in the functional organization of the fetal body and the fetal brain, thereby, possibly inducing alterations in sexuality, including changes in sexual orientation and related cognitive activities (e.g., Joseph et al., 1978; Meyer-Bahlburg, 1993; Reinisch & Sanders, 2012).
Specifically, chronic maternal stress can alter the binding and reception of testosterone, and can prevent testosterone from binding with the Wolfian and Mullerian ducts, and the neurons responsible for sexual behavior. If the fetus is subject to considerable stress a genetic male or female may become homosexual. For example, if "mother" is in an abusive relationship or subject to other profound stress, the secretion of stress-steroids, may effect the sexual differentiation and the development of the the Wolfian and Mullerian ducts as well as the limbic system and induce the female pattern of neural development.
Maternal stress may turn a male into a homosexual, or even a female! This is because stress steroids act to block the reception of androgens, thus inducing the female pattern.
That is, as these stress-steroids, e.g., cortisol and aldosterone, are not true androgens, rather than inducing a male pattern, they may block the reception and binding of fetal testosterone, thereby inducing the female developmental pattern. In fact, the secretion of cortisol and aldosterone--particularly if prolonged--can reduce the secretion of gonadropins (Moberg, 1985), which would also interfere with male sexual behavior and the development of the male pattern of neural development.
LIMBIC SYSTEM SEXUALITY
The limbic system is sexually differentiated such that there is a male, female, and even homosexual limbic system. In humans, sexual differentiation is initiated around 3 months after conception, and is triggered by the presence or absence of testosterone which also effects cellular development. For example, glia cells, which manufacture certain neurotransmitters and which nourish and even guide immature migrating immature neuroblasts to their terminal substrate, develop unique male-specific patterns before birth. Hence, testosterone effects neural migration and thus the organization and neural growth of the limbic system as well as the neocortex and spinal cord.
For example, the presence of fetal testosterone promotes the development of spinal motor neurons which project to the phallus. Moreover, the total brain volume of the human male is about 7% larger than that of the female, and much of this differences is due to the greater volume of white matter in the male cerebrum (glia and axons), the only exceptions being the human hippocampus which is larger in the female, and the amygdala which is 16% larger in the male in total volume (Filipek, et al., 1994).
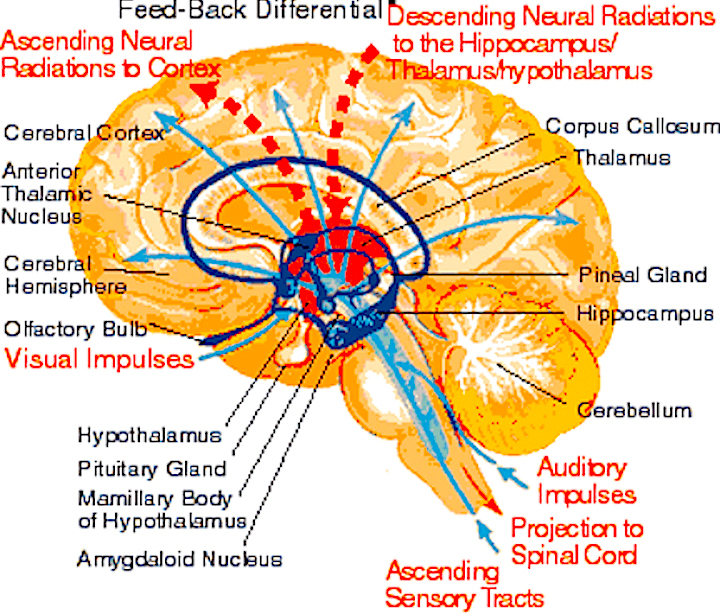
SEXUAL DIMORPHISM IN THE HYPOTHALAMUS
The Sexual Differentiation of the Hypothalamus
As is well known, sexual differentiation is strongly influenced by the presence or absence of gonadal steriod hormones during certain critical periods of prenatal development in many species including humans. Not only are the external genitalia and other physical features sexually differentiated but certain regions of the brain have also been found to be sexually dimorphic and differentially senstitive to steriods, particularly the preoptic area and ventromedial nucleus of the hypothalamus, as well as the amygdala (Bleier et al. 1982; Dorner, 1976; Gorski et al. 1978; Rainbow et al. 1982; Raisman & Field, 1971, 1973).
Indeed it has now been well established that the amygdala and the hypothalamus (specifically the anterior commissure, anterior-preoptic, ventromedial and suprachiasmatic nuclei) are sexually differentiated and have sex specific patterns of neuronal and dendritic development, (Allen et al. 1989; Blier et al. 1982; Gorski et al. 1978; Rainbow et al. 1982; Raisman & Field, 1971, 1973; Swaab & fliers, 1985).
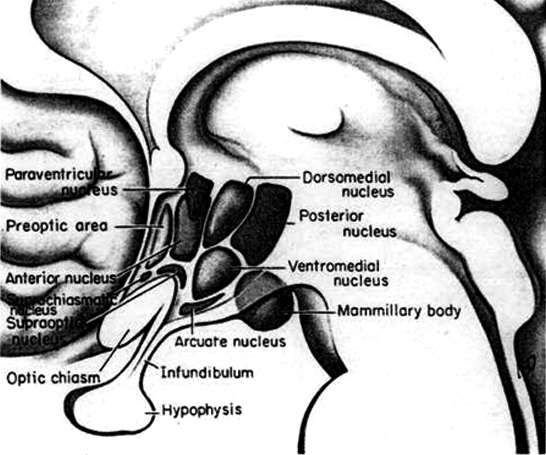
This is a consequence of the presence or absence of testosterone during fetal development in humans, or soon after birth in some species such as rodents. Specifically, the presence or absence of the male hormone, testosterone during this critical neonatal period, directly effects and determines the growth and pattern of interconnections between the amygdala and hypothalamus, between axons and dendrities in these nuclei as well as the hippocampus, septal nuclei, olfactory system (ref), and thus the organization of specific neural circuits. In the absence of testosterone, the female pattern of neuronal development occurs. Indeed, it is the presence or absence of testosterone during these early critical periods that appear to be responsible for neurological alterations which greatly effect sex differences in thinking, sexual orientation, aggression, and cognitive functioning (Barnett & Meck, 1990; Beatty, 2012; Dawson et al. 1975; Harris, 1978; Joseph, et al. 1978; Stewart et al. 1975).
For example, if the testes are removed prior to differentiation, or if a chemical blocker of testosterone is administered thus preventing this hormone from reaching target cells in the limbic system, not only does the female pattern of neuronal development occur, but males so treated behave and process information in a manner similiar to females (e.g., Joseph et al. 1978); i.e. they develop female brains and think and behave in a manner similar to females. Conversely, if females are administered testosterone during this critical period, the male pattern of differentiation and behavior results (see Gerall et al. 2012 for review).
That the preoptic and other hypothalamic regions are sexually dimorphic is not surprising in that it has long been known that this area is extremely important in controlling the basal output of gonadotrophins in females prior to ovulation and is heavily involved in mediating cyclic changes in hormone levels (e.g. FSH, LH, estrogen, progesterone). Chemical and electrical stimulation of the preoptic and ventromedial hypothalamic nuclei also triggers sexual behavior and even sexual posturing in females and males (Hart et al., 1985; Lisk, 1967, 1971) and, in female primates, even maternal behavior (Numan, 1985). In fact, dendritic spine density of ventromedial hypothalamic neurons varies across the estrus cycle (Frankfurt et al., 1990) and thus presumably during pregnancy and while nursing.
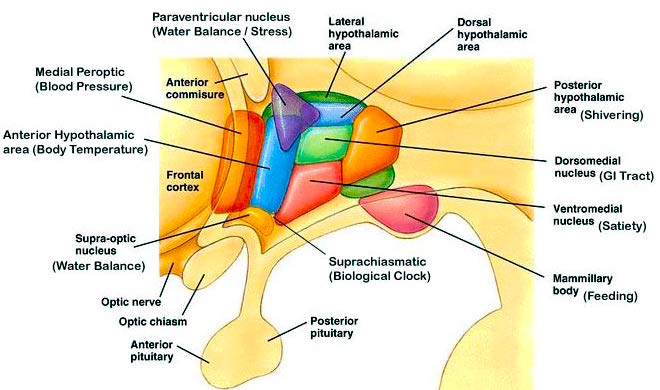
In primates, electrical stimulation of the preoptic area increases sexual behavior in males, and significantly increases the frequency of erections, copulations and ejaculations, we well as pelvic thrusting followed by an explosive discharge of semen even in the absence of a mate (Hart, et al., 1985; Maclean, 1973). Conversely, lesions to the preoptic and posterior hypothalamus eliminates male sexual behavior and results in gonadal atrophy.
Hence, it is thus rather clear than the ability to sexually reproduce is dependent on the functional integrity of the hypothalamus. In fact, it is via the hypothalamus acting on the pituitary, that gonadotropins come to be released. Gonadotropins control the production and/or release of gametes; i.e. ova and sperm.
Specifically, the hypothalamic neurons secrete gonadotropin-releasing hormone, which acts on the anterior lobe of the pituitary which secretes gonadotropins. However, given that in females, this is a cyclic event, whereas in males sperms are constantly reproduced, is further evidence of the sexual dimorphism of the hypothalamus.
Although the etiology of homosexuality remains in question, it has been shown that the ventromedial and anterior nuclei of the hypothalamus of male homosexuals demonstrate the female pattern of development (Levay, 1991; Swaab, 1990). When coupled with the evidence of male vs female and homosexual differences in the anterior commissure which links the temporal lobe and sexually dimorphic amygdala (see below) as well as the similarity between male homosexuals and women in regard to certain cognitive attributes including spatial-perceptual capability (see below), this raises the possibility that male homosexuals are in possession of limbic system that is more "female" than "male" in functional as well as structural orientation.
It is also interesting to note that the sexually dimorphic preoptic region contains thermosensitive neurons, and controls the physiological and behavior responses to excessive external cold or heat. That is, it is responsible for internal thermoregulation and thus heat loss or retention (Alam et al., 1995). Although we can only speculate, it may well be sex differences in this structure which accounts (at least in part) for the stereotypical differences in male vs female perceptions of cold, and why, stereotypically, females (despite their extra-layers of heat-retaining fat) are more likely to insist on elevating room temperature.
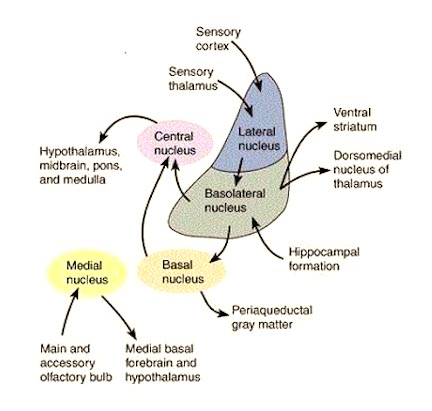
The Sexual Differentiation of the Amygdala
The female and male primate amygdala are sexually differentiated and have their own unique patterns of dendritic growth and organization (Nishizuka & Arai, 1981). As noted, in humans the male amygdala is 16% larger, and in male rats the medial amygdala is 65% larger than the female amygdala (Breedlove & Cooke, 1999), and the male amygdala grows or shrinks in the presence of testosterone--findings which may be related to sex differences in sexuality and aggression.
Moreover, female amygdala neurons are smaller and more numerous, and densely packed than those of the male (Bubenik & Brown, 1973; Nishizuka & Arai, 1981), and smaller, densely packed neurons fire more easily and frequently than larger ones--which may be related to the fact that females are more emotional and more easily frightened than males (chapters 7,13,15), as the amygdala is a principle structure involved in evoking feelings of fear (Davis et al., 1997; Gloor, 1997; LeDoux, 1996).
Dendritic spine density in the female rat hippocampus also increases and decreases by as much as 30% during each estrus cycle (Woolley, et al., 1990) which in turn may influence memory. Indeed, in humans it has been shown that estrogen replacement therapy slows memory loss in women. In fact, it has been reported that women who take this hormone have a 54% lower chance of developing Alzheimers (see chapter 16). On the other hand, dendritic spine density can rapidly change within a few seconds (regardless of gender), as these spines can rapidly grow or disappear in response to varying experiences or lack thereof.
In addition, the human anterior commissure which connects the right and left amygdala/temporal lobe is sexually differentiated (Allen et al. 1989), as is primate/mammalian hypothalamus (Bleier et al. 1982; Dorner, 1976; Gorski et al. 1978; Rainbow et al. 1982; Raisman & Field, 1971, 1973)--with which the amygdala is intimately interconnected. That is, the anterior commissure is thicker in women which, coupled with her more densely packed amygdala neurons (Bubenik & Brown, 1973; Nishizuka & Arai, 1981) may account for her greater social-emotional sensitivity (see chapters 8, 10, 13, 15).
Thus, different structures of the limbic system have sex specific patterns of neuronal and dendritic organization and perform different functions depending on if one is a man or a woman.
For example, chemical and electrical stimulation of the sexually dimorphic preoptic and ventromedial hypothalamic nuclei triggers and/or increases sexual behavior in males and females (with each taking their respective sexual positions), and significantly increases the frequency of erections, copulations and ejaculations, as well as pelvic thrusting followed by an explosive discharge of semen even in the absence of a mate (Hart et al., 1985; Lisk, 1967, 1971; Maclean, 1973). In female primates, activation of these areas can also trigger maternal behavior (Numan, 1985). Conversely, lesions to the preoptic and posterior hypothalamus eliminates male sexual behavior and results in gonadal atrophy. Likewise, activation of the sexually dimorphic amygdala--which is larger in males (Filipek, et al., 1994)-- can produce penile erection and clitoral engorgement (Kling and Brothers, 2012; MacLean, 1990; Robinson and Mishkin, 1968; Stoffels et al., 1980), and trigger sexual feelings (Bancaud et al., 1970; Remillard et al., 1983), extreme pleasure (Olds and Forbes, 1981), memories of sexual intercourse (Gloor, 1986), as well as ovulation, uterine contractions, lactogenetic responses, and orgasm (Backman and Rossel, 1984; Currier, Little, Suess and Andy, 1971; Freemon and Nevis,1969; Warneke, 1976; Remillard et al., 1983; Shealy and Peel, 1957).
Moreover, these sexually dimorphic structures also play different roles among females depending on if a woman is sexually receptive, pregnant, or lactating. For example, in a lactating female, the sexually dimorphic supraoptic and paraventricular hypothalamic nuclei (which projects to the posterior lobe of the pituitary) may trigger the secretion of oxytocin--a chemical which can trigger uterine contractions as well as milk production and which makes nursing a pleasurable experience. In fact, dendritic spine density of ventromedial hypothalamic neurons varies across the estrus cycle (Frankfurt et al., 1990) and thus presumably during pregnancy and while nursing.
Hence, the core of our personal and emotional being, the limbic system, is sexually differentiated. There is a male vs a female limbic system, and even a "homosexual" limbic system (Levay, 1991; Swaab, 1990); structures which are organized in unique sex specific dendritic and neuronal patterns and which govern sex-specific behaviors. Coupled with evolutionary (Joseph, 1999e) and early environmental influences (Joseph, 1979; Joseph & Gallagher, 1980), the sex differences in these and other structures account for many of the stereotypical sex differences in thinking, sexual orientation, aggression, and cognitive functioning (Barnett & Meck, 1990; Beatty, 2012; Dawson et al. 1975; Harris, 1978; Joseph, et al. 1978; Stewart et al. 1975) which characterized the mind of woman and man, including their sexual behaviors.
Stress, Sex Abuse, and Homosexuality
Asignificant number of males with a history of chidhood sexual abuse (CSA) report sexual disturbances (Finkelhor, 1979; Johnson & Shrier, 1985; Rogers & Terry, 1984), including hypo or hypersexuality, compulsive masturbation, and difficulty forming sexual relationships or performing adequately with a female sex partner (Duncan & Williams, 1998; Elliott & Briere, 2012; Hunter, 1991). Some sexually abused males experience considerable confusion over their sexual identity and engage in homosexuality (Johnson and Shrier, 1985; Simari & Baskin, 1984). Finkelhor (1979) reported that males with a history of CSA are four times more likely than non-abused males to engage in homosexual activities. In addition, a significant relationship between CSA and teenage- or adult-onset pedophilia and violent criminal behavior has been reported (Haaspasalo & Kankonon, 1997; Knight & Prentky, 1993; Rubinstein et al., 1993; Watkins and Bentovim, 2012; Widom and Ames, 1994), including violence toward loved ones and intimate partners (Duncan and Williams 1998). As is also the case with women, men who were sexually abused suffer significant emotional problems, including depression, fear, anger, and anxiety, and not infrequently experience homicidal, suicidal, and self-destructive feelings (Mendel 1995; Urquiza and Capra, 1990; Watkins and Bentovim, 2012); emotions, feelings, and behaviors which approximately 30% of those who are "homosexual" also report (see below). Indeed, approximately 30% of male and female homosexuals have significantly alcohol, drug, and serious mental disorders (Family Research Counsel, 1998; Lewis, et al., 1982; Zehner & Lewis, 1985; Ziebold & Mogenson, 1982), and frequently batter their sex partners (Lobel, 1988).
Perhaps not surprisingly, men with a history of CSA, that is, those who have been abused by men rather then women, often become exceedingly homophobic (Urquiza and Capra, 1990). Of those who claim that the "abuse" was a positive experience, these males may well have been homosexual in orientation to begin with (e.g. Johnson and Shrier, 1985).
Homosexuality probably does not have a single "cause." That is, some individuals may become homosexual because of a genetic predisposition (Bailey et al., 1993; Byne & Parsons, 1993), whereas others may engage in these behaviors because they were sexually abused, whereas yet others may have become homosexual due to the profound effects of stress and stress hormones on the limbic system. That is, in these latter instances, the sexual organization and thus the "sexual orientation" of the limbic system may have become altered, and may have assumed the female pattern of sexual differentiation due to profound fetal, neonatal, or childhood stress (e.g., Meyer-Bahlburg, 1993; Reinisch & Sanders, 2012). In fact, because of limbic system sensitivity to circulating stereoids, chronic stress may in fact induce steroidal alterations in the functional organization of the hypothalamus and amygdala, thereby, possibly inducing alterations in sexuality, including changes in sexual orientation and related cognitive activities (e.g., Joseph et al., 1978; Meyer-Bahlburg, 1993; Reinisch & Sanders, 2012).
Specifically, chronic stress can alter the binding of testosterone, and can prevent testosterone from binding with hypothalamic and amygdala neurons responsible for sexual behavior (Raab & Haedenkamp, 1981). If the fetus is subject to considerable stress, for example, if "mother" is in an abusive relationship or subject to other profound stress, the secretion of stress-steroids, may effect the sexual differentiation and the development of the limbic system and induce the female pattern of neural development. That is, as these stress-steroids, e.g., cortisol and aldosterone, are not true androgens, rather than inducing a male pattern, they may block the reception and binding of fetal testosterone, thereby inducing the female developmental pattern. In fact, the secretion of cortisol and aldosterone--particularly if prolonged--can reduce the secretion of gonadropins (Moberg, 1985), which would also interfere with male sexual behavior and the development of the male pattern of neural development.
As noted, it has been shown that the ventromedial and anterior nuclei of the hypothalamus of male homosexuals demonstrate the female pattern of development (Levay, 1991; Swaab, 1990); sex-specific patterns which may have been induced by stress. If these stress induced steroidal changes also negatively impact the amygdala, affected individuals may not only become homosexual, or sexually and emotionally abnormal, but later engage in self-destructive sexual activities including prostitution and indiscriminate homosexuality (see chapter 28).
Corticosteroids when secreted at high levels, can in fact induce wide spread neural injury and abnormal neuroplastic changes throughout the brain. Pyramidal neurons in particular are especially vulnerable to the deleterious effects of stress and high levels of corticosteroids, especially those located in limbic system structures due the abundance of Type II adrenal steroid receptors which abound within this tissue (Lupien & McEwen, 1997; Pugh, Fleshner, & Rudy, 1997). Corticosteroids at high levels can exert a suppressive influences on membrane receptor proteins, thereby altering excitability and information transmission between neurons (Hua & Chen, 1989; Majewska, Harrison, Schwartz, Barker, & Paul, 1986), and can detach the cellular receptor from its attached protein (Beaulieu, 2007); a condition which interferes with messenger RNA protein transcription and thus the genetics of neural differentiation and neural plasticity, including, perhaps, those related to the sexual differentiation of the brain.
Moreover, prolonged, repetitive, profound stress, can induce abnormal activity involving the amygdala (see chapters 28, 29, 30), including seizure-like activity referred to as kindling. Abnormal amygdala (and overlying temporal lobe) activity has been associated with the development of hyposexuality (Taylor, 1971; Heirons and Saunders, 2016; Toon, Edem, Nanjee, and Wheeler, 1989), hypersexuality (Blumer, 1970) as well as homosexuality, transvestism, and thus confusion over sexual orientation (Davies and Morgenstern, 2010; Kolarsky et al., 2017). In fact, abnormal- or seizure activity within the amygdala or overlying temporal lobe may induce an individual to engage in "sexual intercourse" even in the absence of a partner.
Coupled with animal studies which demonstrate that sex specific behaviors and cognitive activities can be enhanced or altered by steroids (Joseph et al., 1978; Reinisch & Sanders, 2012) or suppressed due to early environmental influences, including the stress of deprivation (Joseph, 1979; Joseph and Gallagher, 1980), and given that the sexual differentiation of the hypothalamus may be altered by steroidal manipulations (Raisman and Field, 1973), it can also be assumed that traumatic stress and the massive secretion of steroids, particularly during early sexual development, may alter the neural organization of these same structures. In consequence, sexuality, sexual orientation, as well as emotion, personality, including the ability to cope with stress may be disrupted and become abnormal among those who have been severely or repetitively sexually abused and traumatized.
The Homosexual Limbic System.
As noted above, some men become homosexual or bisexual due to sexual abuse experienced as children (Finkelhor, 1979; Johnson and Shrier, 1985; Simari & Baskin, 1984), whereas in others there is a suggestion of a genetic contribution (Bailey et al., 2013; Byne & Parsons, 1993). Nevertheless, it has been reported that the hypothalamus (i.e. the ventromedial and anterior nuclei) in male homosexuals is organized in a manner similar to the female pattern of development (Levay, 1991; Swaab, 1990). Moreover, the anterior commissure is larger not only in females, but is 35% larger in homosexual males vs male heterosexuals (Allen & Gorski, 2012). Coupled with the evidence reviewed above, this raises the possibility that sex differences in male vs female and male vs homosexual male sexual orientation and thus the capacity to experience sexual pleasure when with a heterosexual or homosexual partner, may be determined by these nuclei.
For example, it could be assumed that homosexual males respond to males with feelings of sexual attraction and desire, as do heterosexual females, i.e. because they are in possession of a "female" limbic system which responds to male physical and facial features with sexual arousal; i.e. the amygdala contains neurons which respond to faces, facial expressions, and which can determine the sex of the individual viewed (e.g. Leonard et al. 1985; Rolls 1984). Conversely, heterosexual males, being in possession of a "male" hypothalamus and amygdala, not only respond to females with sexual arousal, but they behave and act more aggressively than females and homosexual males.
Presumably, it is because homosexuals have a "female" limbic system they they are thus more inclined to behave in a manner similar to women than to men. Indeed, homosexual males (in general) and females tend to be more alike than different in regard to social-emotional reactions and tendencies (Tripp 2007). In some cases these feminine tendencies are grossly exaggerated (Tripp, 2007); i.e. the "swishy" male with the exaggerated high pitched voice.
A significant number of homosexuals, in fact, are psychologically similar to females in a number of ways, including having a high interest in fashion and wearing apparel, a pronounced tendency to employ feminine body language and vocal tones, to shun sports and avoid fights, and to have a fear of physical injury, particularly during childhood (Bell et al. 1981; Bieber et al. 1962; Van Den Aardweg, 2014; Tripp, 2007). Many also tend to maintain intense dependency relations with their mothers and to remain distant from strong male figures including their fathers (Green, 2007); heterosexual males (including male primates; e.g. Fedigan, 2012; Goodall, 1986) tend to behave in a completely different fashion and are far more likely to seek the company of males and men and to engage in rough and tumble activities.
As children homosexual males tended to prefer female companions and friends, girls toys, activities, and often girls clothes, and behaved in an effeminate manner (Bell, et al. 1981; Saghir & Robins, 1973; Grellet et al. 1982; Green, 2007). Indeed, from 67% to 75% of homosexuals vs 2%-3% of heterosexual males reported being "feminine" and more like girls than boys as children (Saghir & Robins, 1973; Green, 2007). Moreover, homosexual males, like heterosexual females, demonstrate comparatively inferior spatial perceptual capabilities as compared to heterosexual males (Gladue et al. 2010; Sanders & Ross-Field, 2006; Wilmot & Brierley, 1984). Homosexuals tend to perform similarly to females on these spatial tests.
It is tempting to speculate that because some homosexuals demonstrate an almost hyper-developed pattern in the structure of the anterior commissure (and thus presumably the amygdala, as well as the hypothalamus), that this may account for why a significant minority of these individuals engage in excessive, dangerous and sometimes "bizarre" (e.g. "fist fucking") sexual behaviors, including indiscriminate promiscuity, "orality," "anality," and a proclivity to engage in group oral/anal sex, or to "cruise" and repeatedly have sex with strangers (sometimes in a single evening). It has been reported that between 24% to 30% of this population have repeatedly engaged in truly reckless and self-destructive sexual behaviors during their youth (Aaron, 1973; Gans, 1993; Pollak, 1993; Symons 1979; Tripp, 2007) --manifestations of the Kluver-Bucy syndrome?
Even at its most benign, male-homosexual sexual-encounters often involve strangers having unprotected sex with strangers. And among younger homosexuals "gay love" is stereotypically often little more than one stranger ejaculating into the anus or the mouth of another stranger, who then swallows or spits, and then goes on his way. Of course many heterosexual males would be happy to ejaculate into the mouth or vagina or a beautiful female stranger. Moreover, like heterosexuals many homosexuals form "long-term" male-male relationships. However, even in so called "committed" homosexual relationships, promiscuity (at least among younger homosexuals) is common (Tripp, 2007).
The hyper-developed pattern of limbic system development, coupled perhaps with abuse/stress-induced abnormalities may also account for the high incidence of emotional disorders including suicide in homosexuals. It has been estimated that approximately 30% of the homosexual population suffer from significant alcohol and drug addiction and/or psychopathology (Family Research Counsel, 2012; Lewis, et al., 1982; Zehner & Lewis, 1985; Ziebold & Mogenson, 1982). Of course, a significant number of homosexuals are exceedingly high functioning, earning six and seven figure incomes, and are devoid of any evidence of pathology or severe emotional disturbances.
Nevertheless, as compared to the general population, severe and profound psychological problems are over represented among homosexuals as a group (Family Research Counsel, 2008; Lewis, et al., 1982; Lobel, 1988; Zehner & Lewis, 1985; Ziebold & Mogenson, 1982), which in turn may be related to possible limbic system abnormalities. This is not to negate the impact of social-environmental influences on the development of these "abnormalities" as the environment can have a significant impact on one's self concept (Joseph 2012b). That is, feeling different, being victimized, rejected, taunted, tormented, isolated, and so on, is in-itself exceedingly stressfull and debilitating, and it would be expected that those victimized in this fashion would experience considerable self-hate, anger, depression, and so on, and would perhaps be inclined to engage in self-destructive behaviors.






