





Rhawn Joseph, Ph.D.
BrainMind.com
SIMPLE, COMPLEX, LOWER- & HIGHER ORDER HYPERCOMPLEX FEATURE DETECTORS
The visual cortex is made up of a variety of cell types each of which is concerned with the analysis of different visual features (Ferster, et al., 1996; Hubel & Wiesel, 1959, 1962, 1968; Kaas & Krubitzer 1991; Sereno et al. 1995). These include simple, complex, and (higher & lower order) hypercomplex cells which are distributed disproportionately throughout areas 17,18, 19.
To briefly summarize, simple cells appear to be involved in the initial analysis of incoming visual cortical input, and are most sensitive to slowly moving stimuli. They are found predominantly within area 17 and in layer IVa,b,c,. Some are sensitive to stimuli moving in one direction, whereas others may respond to stimuli moving in any direction. In fact, almost 95% of the neurons in area 17 are responsive to stimuli moving only in one direction, but not the direction of movement. In addition, simple cells are responsive to the particular position and orientation a stimulus may take. However, for a simple cell to fire, a stimulus must assume a specific orientation and position.
Sinple cells relay this processed information to the far more numerous complex cells which are found predominantly in layers II and III and V, which interact and communicate with one another including with layer IV which receives thalamic input. Each complex cell receives input from several simple cells. Complex cells are also concerned with orientation of the stimulus. However, these cells are more flexible and will respond and analyze a stimulus regardless of its particular orientation. These cells via the combined input from simple cells, are probably involved in the earliest stages of actual form perception, i.e. the determination of the outline of an object. A considerable number of complex cells receive converging input from both eyes, the remainder being monocular. Complex cells are found predominantly within area 18.
Hypercomplex cells are concerned with the analysis of discontinuity, angles and corners, as well as movement, position, and orientation. That is, these cells respond selectively to certain visual configurations and thus act so as to determine precise geometric form. It is also via the action of these cells (in conjuction with visual neurons in the temporal lobe) that the first stages of visual closure are initiated. This in part requires that the functional activity of these cells be suppressed such that when presented with an incomplete figure these cells are overridden and the brain is able to "fill in the gaps" in stimuli perceived. It is also for this reason that one does not notice his or her "blind spot"; it is filled in. Hypercomplex cells are found predominantly within area 19.
ASSOCIATION AREAS 18 & 19
Areas 18 and 19 are are involved in the translation and interpretations of visual impressions transmitted from area 17. Although simple and complex cells are found in the association cortex, this region is predominantly populated by hypercomplex (both higher and lower order) neurons --most of which are concerned with the determination of precise geometric form as well as the assimilation of signals transmitted from the primary cortex (Kaas & Krubitzer 1991).
[-INSERT FIGURE 10 ABOUT HERE-]
In contrast to the neurons within area 17, many of the cells within area 18 receive binocular input and can be activated by either eye (Hubel & Wiesel, 1970). This same pattern of bincularity is evident in the parietal association area. It is via the action of these cells that one is able to gather information regarding distance, discrepancies in stimulus location and thus determine depth and achieve stereoscopic vision (Blakemore, 1970a; Hubel & Wiesel, 1970a). Indeed, some association neurons will only fire when a target is a definite distance from the eye.
Many of the neurons in this region, particularly area 19, receive higher order converging input from the parietal and temporal lobe. For example, in addition to visual input, neurons in the superior portions of area 19 respond to tactile and proprioceptive stimuli, whereas those in the inferior portions respond to auditory signals (Morrell, 1967). It is probably in this manner (in conjunction with subcortical connections with, for example, the superior colliculus) that one is able to orient toward and gaze upon an auditory stimulus as well as maintain stabilization of the head (via proprioceptive-vestbibular input) while engaged in visual search.
Hence, overall, the visual association area appears to be involved in the initial analysis of form, distance, and depth perception, as well as the performance of visual closure. It is thus heavily involved in the association of various visual attributes so that a variety of qualities may be ascertained. This would include an objects shape, length, thickness, and color (Sereno et al. 1995).
THE TEMPORAL/PARIETAL VISUAL AREAS
The visual association areas also maintain intimate relationships with the parietal visual regions (area 7) as well as the visual areas in the middle and inferior temporal lobes (ares 37) including V5 (the lateral occipital-temporal junction). The temporal visual areas are in turn reciprocally interconnected with area 7 and areas 17, 18, 19.
As noted, whereas the temporal lobes perform complex form recognition and central and upper visual field analysis, the parietal lobes observe the periphery and lower visual field (where the hands and feet are most likely to be viewed) and together these cortical regions compute and make possible, eye-hand, or hand-object coordination. Hence, a complex interactional visual loop is maintained.
For example, the inferior-medial temporal lobe is concerned with form perception and the analysis of emotional-motivational significance and transmits this information to area 7. Area 7 is involved in visual attention, visual fixation, and the analysis of distance, depth, and objects within grasping distance. Area V5 (at the temporal occipital border) perceive motion. Hence, through this temporal/occipital/parietal pathway, the individual can recognize and fixate on the object (temporal lobe), determine the speed at which it is traveling (V5) and reach out and grasp it (parietal lobe).
As noted, the temporal parietal visual areas are also interconnected with areas 17, 18, and 19, and receive their own autonomous visual input, via, for example, the pulvinar and midbrain. Moreover, the temporal parietal visual areas receive some visual signals in advance of the primary and association visual areas. Area V5, for example, perceive rapid movement, and if injured or deactivated, the ability to perceive rapid movement is diminished or abolished, whereas slow motion perception is maintained due to preservation of the occipital visual areas (Zeki, 1997). Moreover, fast moving stimuli are perceived and activate area V5 in advance of area 17, which in turn is excited by slow moving stimuli in advance of area V5. Hence, V5 receives information from the retina regarding fast moving objects, information that bypasses the primary visual area (Zeki, 1997); a very adaptive relationship, at least from an evolutionary and life-preservative perspective, for under some circumstances it would be most advantageous to perceive something that was moving rapidly, such as a predator, than something less dangerous that was moving slowly.
MULTIPLE VISUAL REALITIES
As detailed in chapters 2, 10, the brain is not only interactional, but is characterized by normal discontinuities where some areas do not always communicate together efficiently. That is, the functioning of the brain is not always in parallel, such that some areas function in semi-isolation or semi-independently giving rise to multiple streams of conscious awareness which may not always correspond to reality. The same can be said even regarding those areas of the brain which are ostensibly concerned with the same visual stimulus, such as auditory or vision as is the case with V5 and areas 17, such that when these stimuli are processed, an internal reality which is slightly different from external reality may be produced.
According to Zeki (1997, pp. 170-171) the evidence has "led us to the concept of dynamic parallelism, by which we mean that while both areas can be healthy and functional in a normal brain, which area gets activated first depends on the nature of the stimulus." However, as also pointed out by Zeki (1997), even within the visual system the various "perceptual systems are therefore different as are the processing systems" and "that there is no synchronizer in the brain capable of setting the results of the operations of the two processing-perceptual systems to time zero... that we thought was the hallmark of our visual experience.... the brain, therefore missychronizes and misbinds or rather more accurately, it binds the results of its processing-perceptual systems, not what happens in real time in the real world."
HOMONYMOUS HEMIANOPSIA & QUADRANTANOPSIA
Massive unilateral destruction of the visual cortices results in blindness in the contralateral temporal visual field and ipsilateral nasal visual field. Hence, a left visual cortex lesion produced a right homonymous hemianopsia. However, these visual disturbances may also result from destruction of the optic radiations or optic tract. It is noteworthy that patients are often unaware of having lost half their sight, particularly with right occipital lesions. Nevertheless, patients may complain of bumping into people and objects they cannot see.
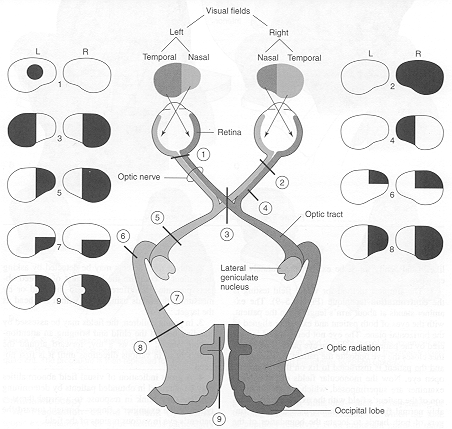
As noted, above, the parietal lobes are concerned with the lower visual fields, whereas the temporal lobes receive massive upper visual field projections. Thus, lesions to the inferior or superior occipital lobe can result in an upper or lower visual field homonymous quadrantanopsia.
HALLUCINATIONS
Electrical stimulation of or lesions involving areas 18 and 19 can produce complex visual hallucinations (Foerster, 1929, cited by Brodal, 1981 & Hecaen & Albert, 1978; Tarachow, 1941), such as images of men, animals, various objects and geometric figures, liliputian-type individuals, including micropsias and macropsias (see Luria, 1980; & Hecaen & Albert, 1978, for review). Sometimes objects may seem to become telescoped and far away, wheras in other situations, when approached, objects may seem to loom and become exceedingly large.
Complex hallucinations are usually quite vivid and fully formed and the patient may think what he sees is a real (Hecaen & Albert, 1978). Foerester (1928, cited by Heacen & Albert, 1978) reported a ptient who hallucinated a butterfly then attempted to catch it when area 19 was elecrically stimulated. Another hallucinated a dog and then called to it, denying the possibility that it was not real.
Complex hallucinations, although usually associated with tumors or abnormal activation of the visual association area, have also been reported with parietal-occipital involvement (Russell & Whitty, 1955), occipital-temporal, or inferior-temporal damage (Mullan & Penfield, 1959; Tarachow, 1941; Teuber et al., 1960), or with lesions of the occipital pole and convexity (Hecaen & Albert, 1978)
Laterality.
According to Hecaen and Albert (1978) based on their review of the international literature, although simple hallucinations are likely following damage to either hemisphere, complex hallucinations are usually associated with right rather than left cerebral lesions (Teuber et al., 1960; Mullan & Penfield, 1959; Hecaen & Albert, 1978).










